重庆巫山大溪遗址动物骨骼鉴定报告
一、遗址概况
大溪遗址位于渝东北的巫山县大溪乡大溪村二社和三社(2001年后合并为三社),遗址所在地的小地名当地习称上溪。遗址的中心地理坐标为北纬31°0′15″,东经109°30′31″,海拔125~145米,高于长江常年枯水位约35~45米。东距巫山县城水路里程37千米、公路里程45千米,西距奉节县15千米。为配合三峡库区地下文物的考古发掘工作,重庆市博物馆考古队(2000年底以考古队为基础成立重庆市文物考古所,2005年底独立运行,2011年更名为重庆市文化遗产研究院,2021年更名为重庆市文物考古研究院)承担了巫山县大溪遗址的考古发掘任务,并组建了大溪遗址考古工作队,于2000~2003年对大溪遗址进行了抢救性考古发掘,发掘总面积共计超过9000平方米。遗迹主要以墓葬、灰坑为主,此外还包括窑、灶、沟、房址等,其中墓葬共计197座、灰坑815个。时代囊括新石器时代一至六期:一至四期为大溪文化(四期含少量玉溪上层文化、玉溪坪文化早期因素)、五期为玉溪坪文化遗存(含部分从下层扰动上来的大溪文化器物);六期为中坝文化老关庙期遗存,商周,汉至六朝,唐宋,明清时期,另外还存在次生堆积(主要是1975年10月至1976年1月时发掘的墓葬区回填后又被长江洪水冲毁后,混合了部分垮塌的原生堆积)[1]。在对骨骼所在的遗迹单位进行登记时,我们发现有个别单位并未进行分期,因此单列为“其他新石器时期”。在对这批骨骼进行整理的过程中,我们按照文化的分期和时代,分别对动物骨骼进行数量的统计以及种属的鉴定。
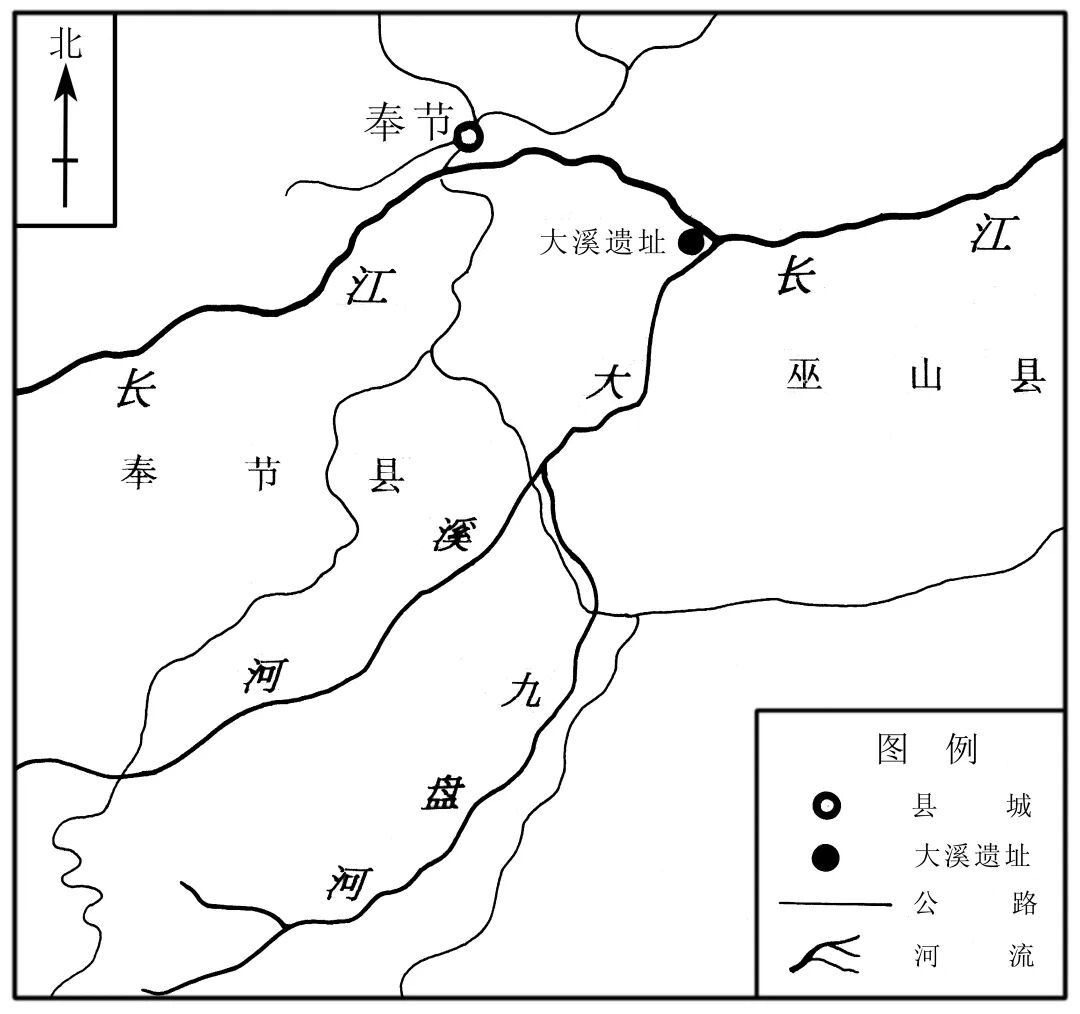
图一 大溪遗址位置图
二、动物数量统计
(一)种类统计
2014年3月~6月,重庆市文物考古研究院动物考古实验室对大溪遗址出土的所有动物骨骼开展了鉴定工作。遗址出土的动物骨骼保存状况整体较好,在埋藏过程中受水流搬运、日晒风化、挤压变形等的影响并不大;发掘时间虽早,但是发掘后及时装箱保存,将对骨骼的破坏降到了最低。
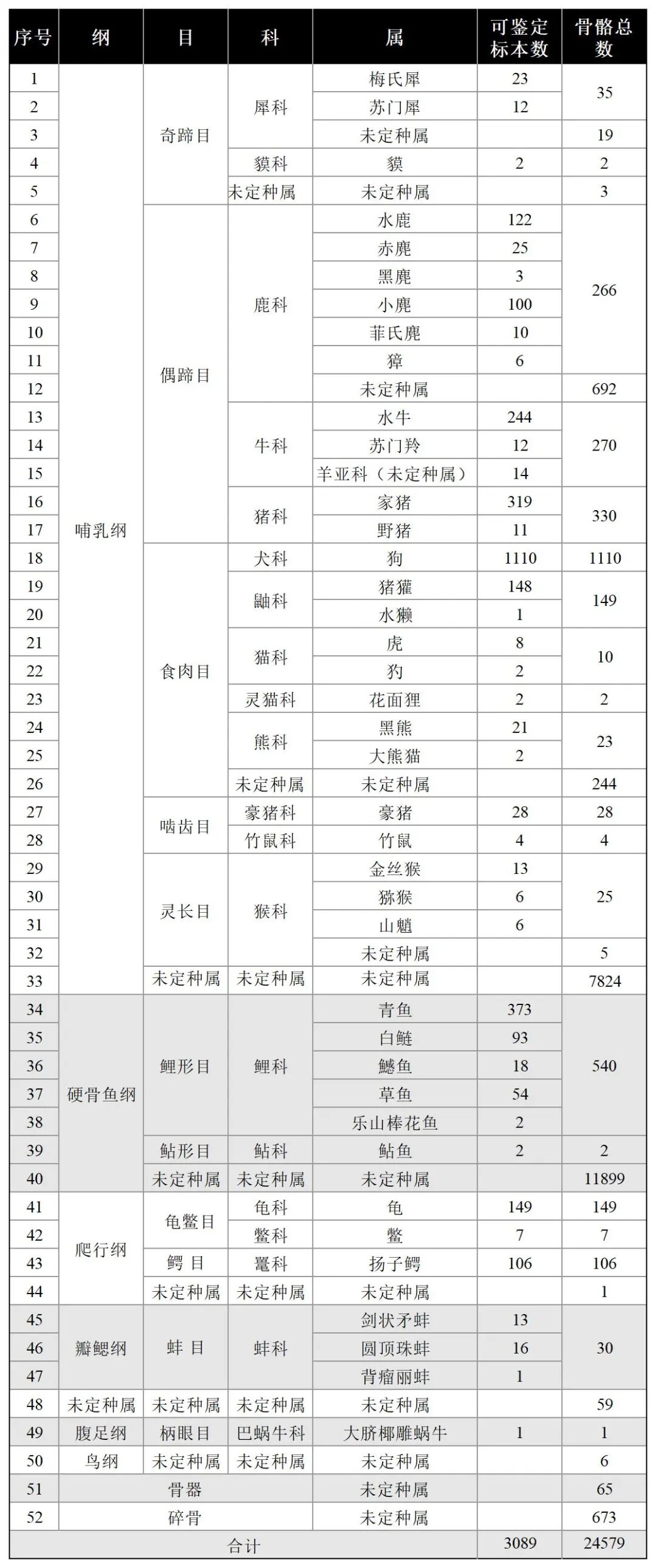
本次共鉴定出的动物种类有哺乳纲、硬骨鱼纲、爬行纲、瓣鳃纲、鸟纲5大类共计40种动物。其中哺乳纲包括奇蹄目3种:犀科梅氏犀、苏门犀,貘科貘;偶蹄目11种:鹿科水鹿、赤麂、黑麂、小麂、菲氏麂、獐,牛科水牛、苏门羚,羊亚科(未定种属),猪科家猪、野猪;食肉目8种:犬科狗,鼬科猪獾、水獭,猫科虎、豹,灵猫科花面狸,熊科黑熊、大熊猫;啮齿目2种:豪猪科豪猪,竹鼠科竹鼠;灵长目3种:猴科金丝猴、猕猴、山魈。硬骨鱼纲鲤形目5种:鲤科青鱼、白鲢、鳡鱼、草鱼、乐山棒花鱼;鲇形目1种:鲇科鲇鱼。爬行纲类龟鳖目2种:龟科乌龟、鼋科鳖;鳄目1种:鼍科扬子鳄。瓣鳃纲蚌目3种:蚌科剑状矛蚌、圆顶珠蚌、背瘤丽蚌;柄眼目1种:大脐椰雕蜗牛。此外,还包括未定种属鸟纲。出土动物骨骼共计24579件,除骨器65件、碎骨673件外,哺乳纲骨骼共计11041件,可鉴定标本数2254件;硬骨鱼纲骨骼共计12441件,可鉴定标本数542件;爬行类263件,可鉴定标本数262件;瓣鳃纲类骨骼共计90件,可鉴定标本数31件;鸟纲骨骼总计6件(表一)。
表一 大溪遗址出土动物骨骼数量统计表
需要说明的是:(1)对未分期的单位,另行列出进行统计;(2)为了避免与遗址中出土的其他遗物重复编号,在所有骨骼编号前均以B(标本)开头另行编号,如2000WDⅡH96:B498;(3)有个别骨骼在发掘时已进行编号,以发掘时的编号为准。
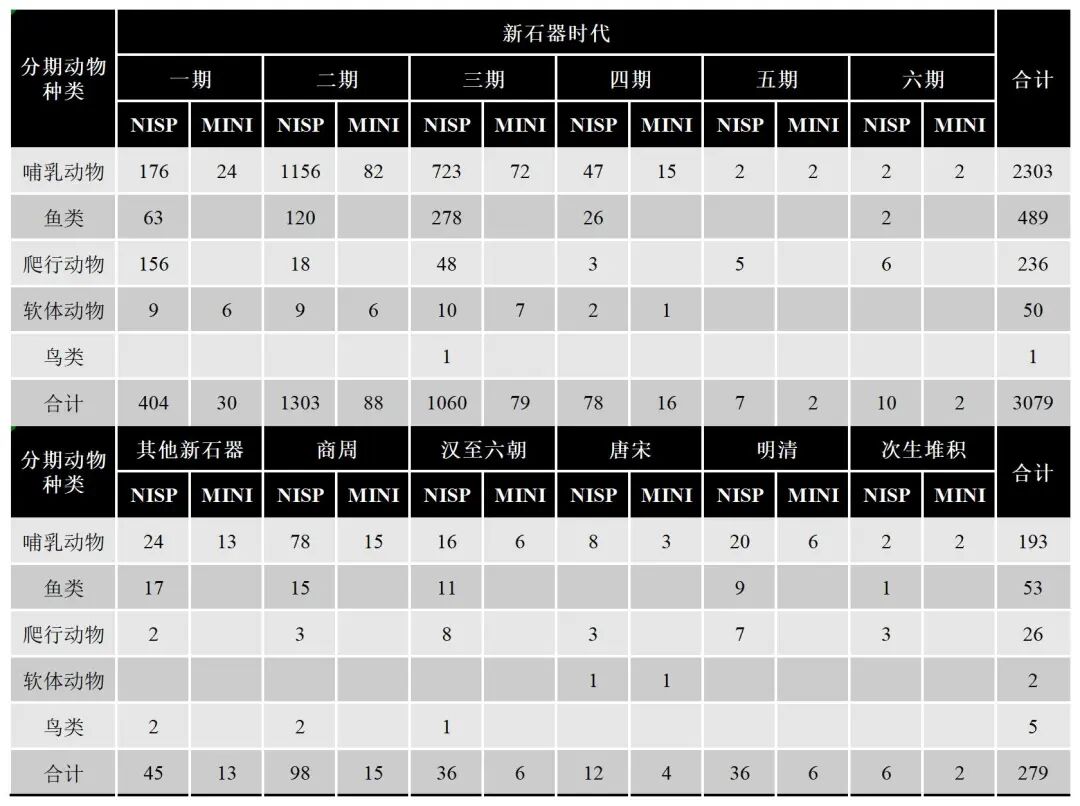
(二)分期统计
分期统计情况见表二、附表一和附表二。
一期(大溪文化一期)哺乳动物共计17种,可鉴定标本数176件,最小个体数23;鱼类2种,可鉴定标本数63件;爬行动物3种,可鉴定标本数156件;软体动2种,可鉴定标本数9件,最小个体数6。
二期(大溪文化二期)哺乳动物共计23种,可鉴定标本数1156件,最小个体数80;鱼类共计4种,可鉴定标本数120件;爬行动物3种,可鉴定标本数18件,软体动物3种,可鉴定标本数9件,最小个体数为6。
三期(大溪文化三期)哺乳纲种类共计 23种,可鉴定标本数 723件,最小个体数 73;硬骨鱼纲共计 6种,可鉴定标本数 278件;爬行纲 2种,可鉴定标本数 48件;瓣鳃纲 3种,可鉴定标本数 10件,最小个体数 7;鸟纲未定种属,可鉴定标本数 1件。
四期(大溪文化四期)哺乳动物共计11种,可鉴定标本数47件,最小个体数15;鱼类共计3种,可鉴定标本数26件;爬行动物共计2种,可鉴定标本数3件;软体动物1种,可鉴定标本数2件,最小个体数1。
五期(玉溪坪文化)出土哺乳动物共计2种,可鉴定标本数2件,最小个体数为2;爬行动物2种,可鉴定标本数5件。
六期(中坝文化老关庙期)出土哺乳动物共计2种,可鉴定标本数为2,最小个体数为2;鱼类1种,可鉴定标本数2件;爬行动物2种,可鉴定标本数6件。
其他新石器时代哺乳动物种类11种,可鉴定标本数为24件,最小个体数13,鸟类未定种属,可鉴定标本数2件;鱼类2类,可鉴定标本数17;爬行动物1种,可鉴定标本数为2件。
商周时期哺乳纲种类共计10种,可鉴定标本数78件,最小个体数15;硬骨鱼纲1种,可鉴定标本数为15件;爬行纲2种,可鉴定标本数3件;鸟纲未定种属,可鉴定标本数2件。
汉至六朝时期出土哺乳纲种类共计5种,可鉴定标本数16件,最小个体数6;硬骨鱼纲共计3种,可鉴定标本数11件;爬行纲共计2种,可鉴定标本数8件;鸟纲未定种属,可鉴定标本数1件。
唐宋时期的哺乳动物共计3种,可鉴定标本数为8件,最小个体数为3;爬行动物1种,可鉴定标本数为3件;软体动物1种,可鉴定标本数为1件,最小个体数1。
明清时期哺乳动物种类共计6种,可鉴定标本数20件,最小个体数为6;鱼类共计2种,可鉴定标本数9件;爬行动物1种,可鉴定标本数为7件。
次生堆积出土的哺乳动物种类共计2种,可鉴定标本数2件,最小个体数为2;鱼类1种,可鉴定标本数为1件;爬行动物1种,可鉴定标本数为3件。
表二 大溪遗址各时期出土动物骨骼可鉴定标本数与最小个体数统计表
三、动物种类描述
(一)哺乳纲 Mammalia
1.奇蹄目Perissodactyla
(1)犀科Rhinocerotidae
出土犀牛骨骼共计53件,主要有臼齿、肩胛骨、掌/跖骨、趾骨等,其中不能鉴定到种的门齿1件、臼齿2件、肩胛骨4件、掌/跖骨5件、指/趾骨7件,时代包括一至三期、其他新石器时代、商周。苏门犀和梅氏犀骨骼共计35件。
梅氏犀 Stephanorhinus kirchbergensis(Jäeger, 1839)
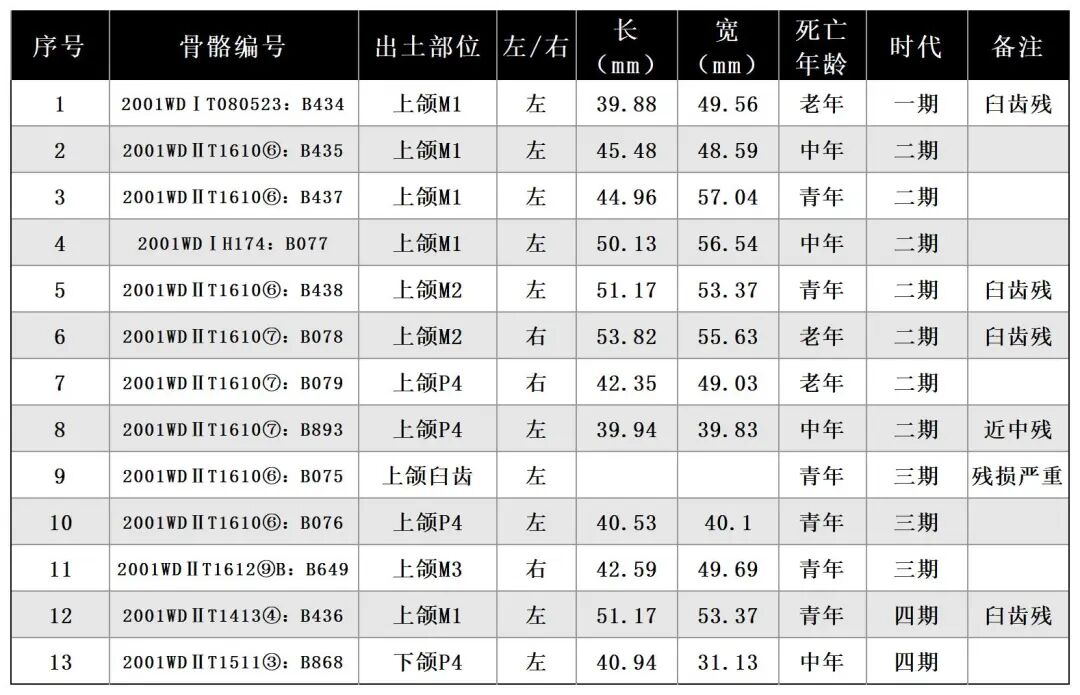
骨骼分布部位及数量:臼齿13枚(表三)、掌/跖骨8件(表四),跟骨1件,时代为一至五期。可鉴定标本数23,最小个体数8。
2001WDⅡT1511③:B868,左侧下颌骨,时代为四期。保存较差,残留一小段水平支,仅保留P4一枚臼齿,颊侧近中面的牙齿有缺损。齿冠较高,臼齿被分为前叶和后叶两部分,后叶比前叶宽;后谷明显较前谷开阔;颊侧凹更深,从暴露出的齿根判断,应有2个齿根。磨损较严重,牙釉质明显高于水泥质,水泥质内凹,从牙齿磨损程度分析,该个体的死亡年龄应处于中年阶段。全长76.64、臼齿高72.54、长×宽:40.94×31.13毫米(图二,1)。
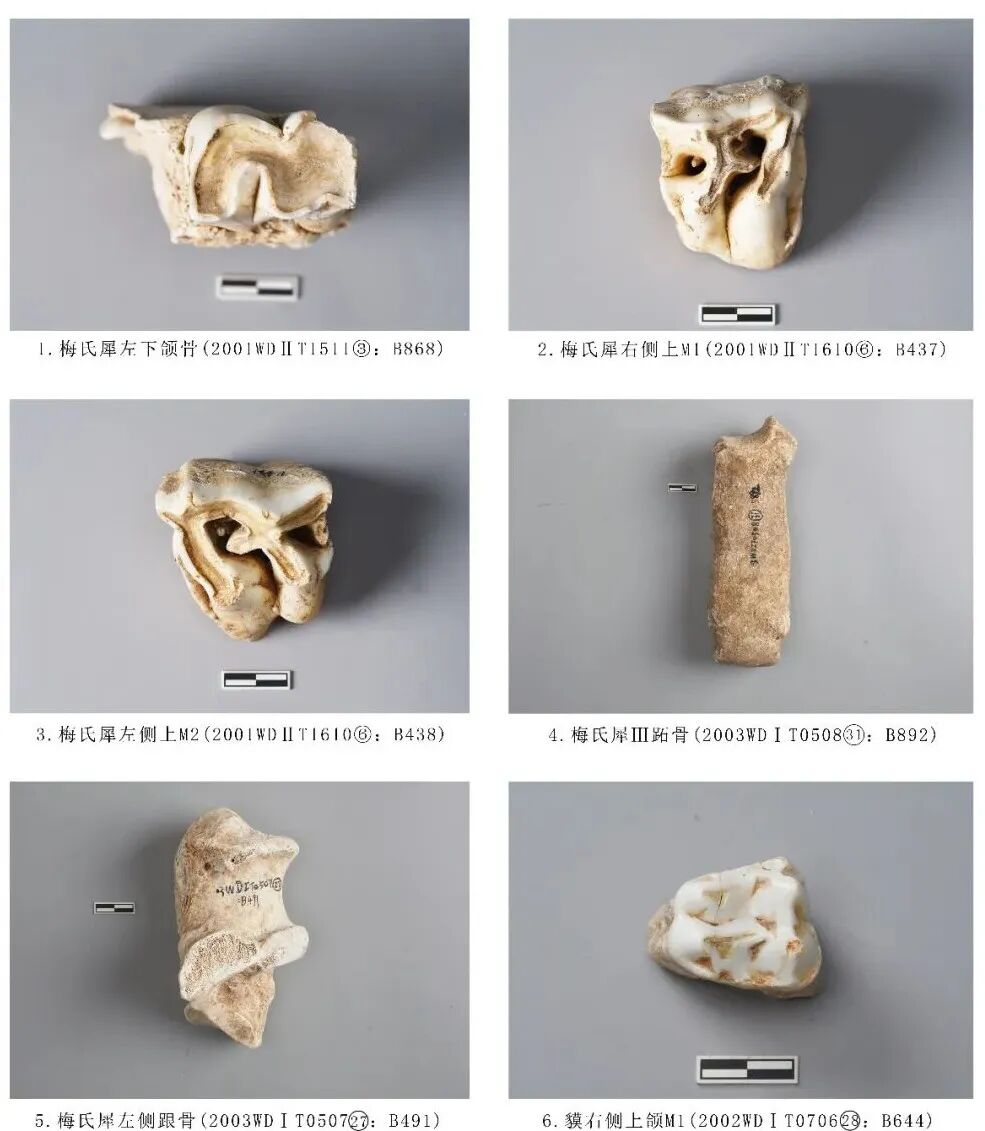
图二 犀科、貘科动物骨骼
2001WDⅡT1610⑥:B437,右侧上颌M1,时代为二期。齿根缺齿,保存臼齿牙釉质部分。齿冠整体呈梯形,宽大于长,底座宽大,向上斜收。前附尖发育较差;后附尖极短而宽;前尖发育程度明显高于前附尖;原尖宽大,前、后收缩明显,有残损;后尖在各尖之中为最大;次尖小,前、后收缩明显,有残损。原脊与外脊的夹角在 60°-70° 之间,内半部膨大;外脊宽阔,其上有一微弱小刺;后脊窄,具粗壮的前刺。中窝不封闭;中谷开口为“V ”型;后窝小。齿带在前、后两侧存在,前齿带强壮,后齿带封闭后窝[2]。臼齿冠面齿脊呈带状磨损,远中面齿脊为暴露出水泥质,从牙齿磨损程度分析应为青年阶段。臼齿残高56.6毫米(图二,2)。
2001WDⅡT1610⑥:B438,左侧上颌M2,时代为二期。齿根缺失,仅保存齿冠部分。M2整体结构与M1相似,齿冠整体呈梯形,颊侧宽于舌侧;附尖发育,前尖褶肋明显,但向下延伸不远;小刺发育,中谷开口成“V”型,比M1窄;后窝很大并且很深;原尖收缩强烈,远中齿带不发育[3]。残高50.93毫米(图二,3)。
表三 梅氏犀臼齿测量数据一览表
2003WDⅠT050831:B892,时代为一期。左侧第Ⅲ掌/跖骨。保存完整。骨骼颜色整体呈灰黑色,推测是烟熏导致。整体宽扁,外廓整体略呈亚腰型,近中关节平面呈“凸”字型,前面观内侧向上斜收形成弯曲弧度;滑车矢状脊前侧不发育,后侧位于中间部位,后侧左右并排两个较深的窝(图二,4)。
表四 梅氏犀掌/跖骨测量数据一览表
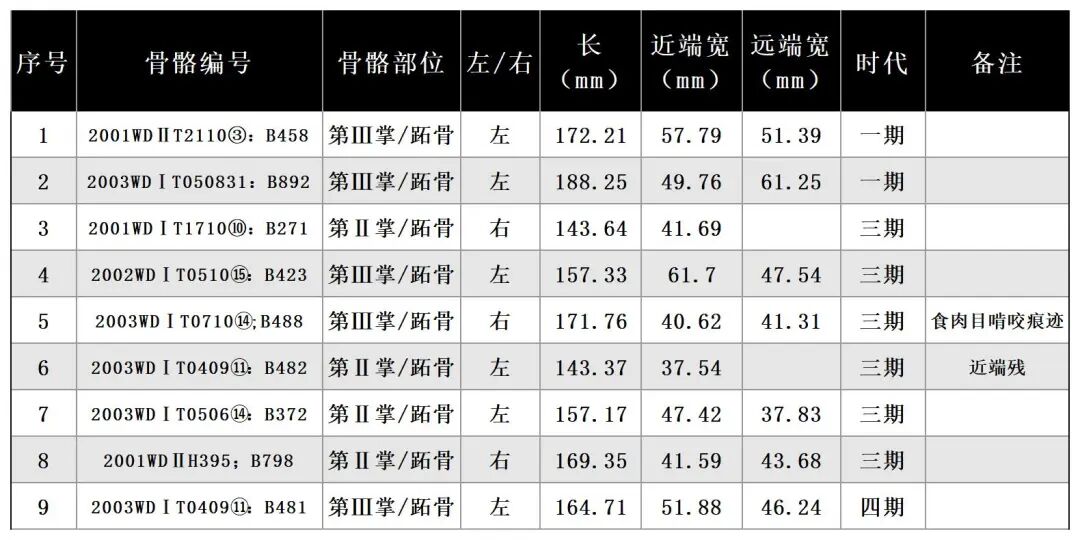
2003WDⅠT050727:B491,左侧跟骨,时代为二期。保存较为完整,载距突右部分缺失。骨骼整体粗壮,近端关节面有一明显的锥形隆起,跟骨体上下两端均突出形成亚腰形;跟骨前突截缘面宽大。全长126.89、近端关节宽65.16,远端最大宽71.19毫米(图二,5)。
苏门犀Dicerorhinus sumatrensis
骨骼分布部位主要为掌/跖骨,时代包括一至三期。可鉴定标本数为12,最小个体数4。
2000WDⅡH96:B498,左侧第Ⅲ掌/跖骨,时代为二期。保存较好,仅在远端滑车上方两侧有残损。近端关节面呈“凸”字型,较平,内侧斜收弧度微弱;远端关节面两侧往中间收,呈骨干宽、关节面窄的状态,滑车矢状脊前侧不发育,后侧位于中间部位。骨干前侧中部位置分布有明显的砍砸痕迹[4]。
(2)貘科Tapiridae Burnett
貘Tapirus sinensis owen
出土牙齿2枚,时代为二、三期。最小个体数为2。
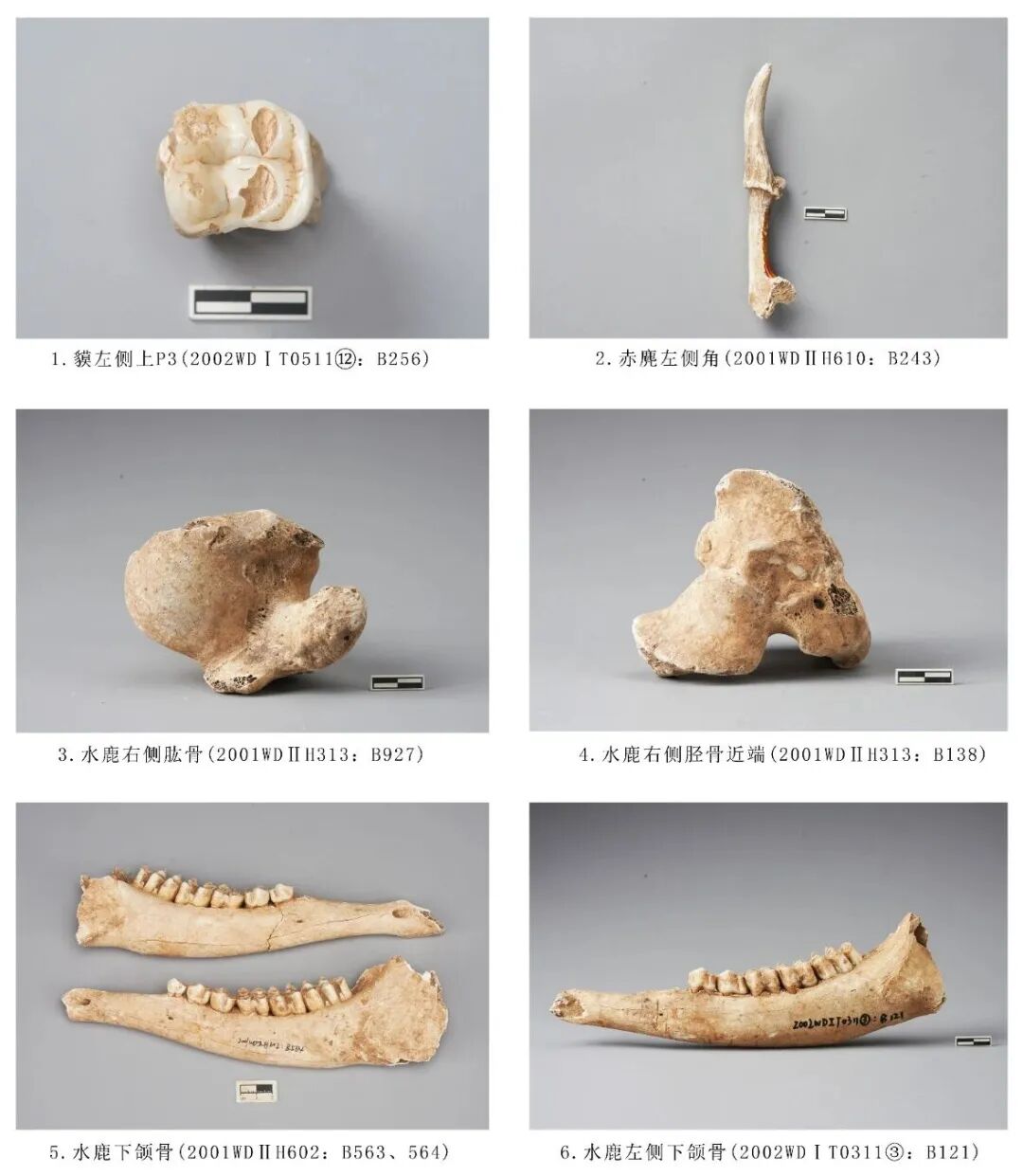
2002WDⅠT070628:B644,右侧上颌M1,时代为二期。保存较好,齿根部分骨皮有脱落。釉质层较厚,齿冠近中远中面均有明显的接触面,保留3齿根(颊侧2,舌侧1)。齿冠面近似梯形,颊侧宽、舌侧窄;颊侧前附尖、前尖、后尖、后附尖相连成脊,前尖>后尖>前附尖>后附尖,4尖相连成脊形成3个齿沟,近中侧齿沟不明显,中间齿沟最宽大,呈“V”形 ;舌侧齿尖粗壮,原尖>次尖,2尖之间有深而宽的齿沟;前尖与原尖形成齿脊,且前附尖、前尖与原尖形成一个宽大的内凹面,高度高于后尖、次尖相连成脊形成的内凹面;颊侧近中面发育有齿带。舌侧2齿尖暴露片状齿质点。从牙齿磨损程度分析该个体的死亡年龄应处于青年阶段。臼齿长31.62、颊侧宽27.53、舌侧宽22.60毫米(图二,6)。
2002WDⅠT0511⑫:B256,左侧上颌P3,时代为三期。颊侧远中缘齿冠有崩残。有3齿根,舌侧的1齿根最为粗壮。牙齿整体成矩形,共有4个齿根:颊侧前尖高大、粗壮,后尖残损;舌侧原尖、次尖磨损内凹,暴露齿质,原脊和后脊几乎平行,中脊开阔。臼齿长×宽:25.28×23.16毫米(图三,1)。
图三 貘科、鹿科动物
2.偶蹄目
(1)鹿科 Cervidae
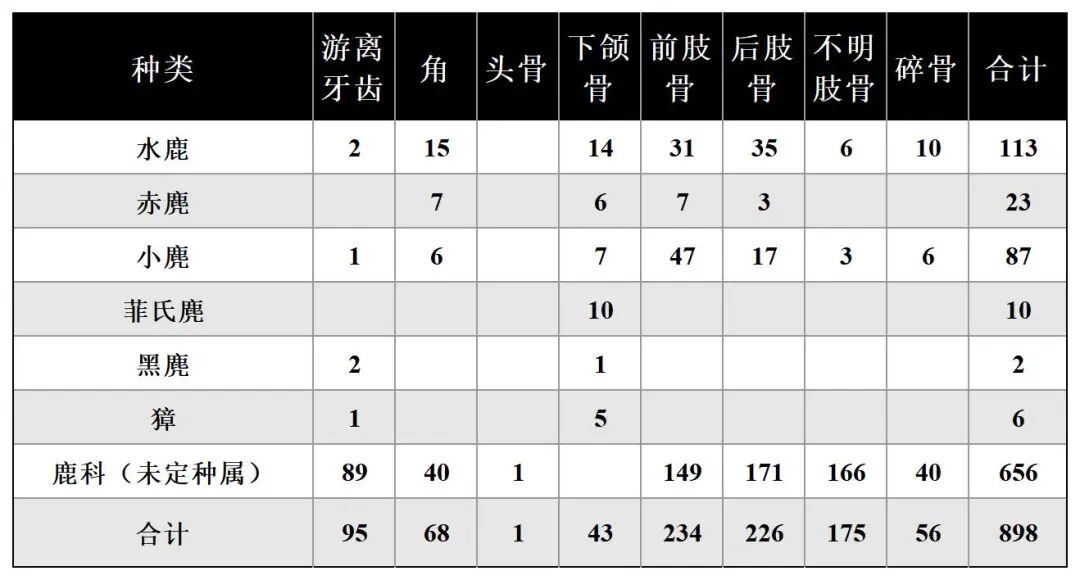
鹿科动物骨骼总计958件,可鉴定标本数为266件。动物种类有水鹿(46%)、赤麂(9%)、小麂(38%)、菲氏麂(4%)、獐(2%)、黑麂(1%)6种。而新石器时代所占数量最多,骨骼总量为898件,可鉴定标本数为242件。骨骼部位包括游离牙齿、角、头骨、下颌骨、肢骨等(表五)。
表五 新石器时期鹿科动物骨骼出土数量统计表
水鹿Rusa unicolor
可鉴定标本数122件,最小个体数17。时代包括一至五期、其他新石器时期、商周、明清。出土的骨骼部位包括游离牙齿、角、下颌骨、肢骨等。
2000WDⅠT050220:B292,右角角柄,时代为四期。保存较差,角心位置缺失,端口深浅不一,无法观察其折断原因,近中处断面新鲜且平滑,应是发掘所致。在角心位置断裂处远中面有一穿孔,与角心中部断裂处相通。靠近头骨部位有明显的烧灼痕迹,且有的部分颜色发灰、发白。残长143.7毫米(图九,3)。
2001WDⅡH602:B564,左侧下颌骨,时代为二期。保存完整的水平支以及全套臼齿,上升支大部分缺失。M1~M3均发育有齿柱,且M1的齿柱有磨损。齿冠面的珐琅质明显高于水泥质,且中部的新月形脊呈颊侧低舌侧高的形态。从牙齿磨损程度分析,该个体的死亡年龄应处于中年阶段。残长243.17、臼齿齿列长112.1毫米,M1~M3长70.85毫米(图三,5)。
2002 WDⅠT0311③:B121,左侧下颌骨,保存完整水平支,P2缺失,保留齿槽,附着P3~M3五枚臼齿。5枚臼齿磨损均暴露出水泥质,新月形脊在M1~M3明显。从牙齿磨损程度分析,该个体的死亡年龄应处于中年阶段。下颌骨残长243.12、臼齿齿列长108.23毫米(图三,6)。
2001WDⅡH313:B927,右侧肱骨,时代为三期。残,仅保留近端关节部分。大结节明显突起,小结节处残损,肱骨头平滑,三角肌粗隆突出。残长135.92、近端最大宽76.69毫米(图三,3)。
2001WDⅡH313:B138,右侧胫骨,时代为三期。残,仅保留近端关节部分。外髁、内髁呈不规则三角形,肌腱沟夹角呈圆角锐角,胫骨粗隆呈瘦长三角形,近端后侧两条腘肌线顺延。残长96.09、近端关节最大宽90.41毫米(图三,4)。
赤麂Muntiacus muntjak
可鉴定标本数为25件,最小个体数11。时代包括一至四期、商周、汉至六朝,骨骼分布部位有角、下颌骨、肢骨、掌跖骨等。
2001WDⅡH610:B243,左侧角,时代为二期。角冠远端有风化脱皮现象,略残,附带小部分额部骨骼。角柄粗长,角环明显突起,角环内侧面生长一突起的分叉,但不延伸,角冠主叉向内向后倾斜,布满纵向生长纹路。残长128.85、角冠残长68.69毫米(图三,2)。
2003WDⅠT050614:B371,右侧下颌骨,时代为三期。水平支保存完整,门齿缺失,附着全套臼齿(P2~M3),上升支残损。下颌骨整体较纤细,下颏孔位置靠前,贴近门齿齿槽位置。臼齿磨损痕迹较重,水泥质内凹,明显低于牙釉质,以P3后半部至M3磨损最为严重,M1~M3发育齿柱,已磨损。推测其死亡年龄应处于中年阶段。残长156.8、齿列长75、M1~M3长44.7毫米(图四,6)。
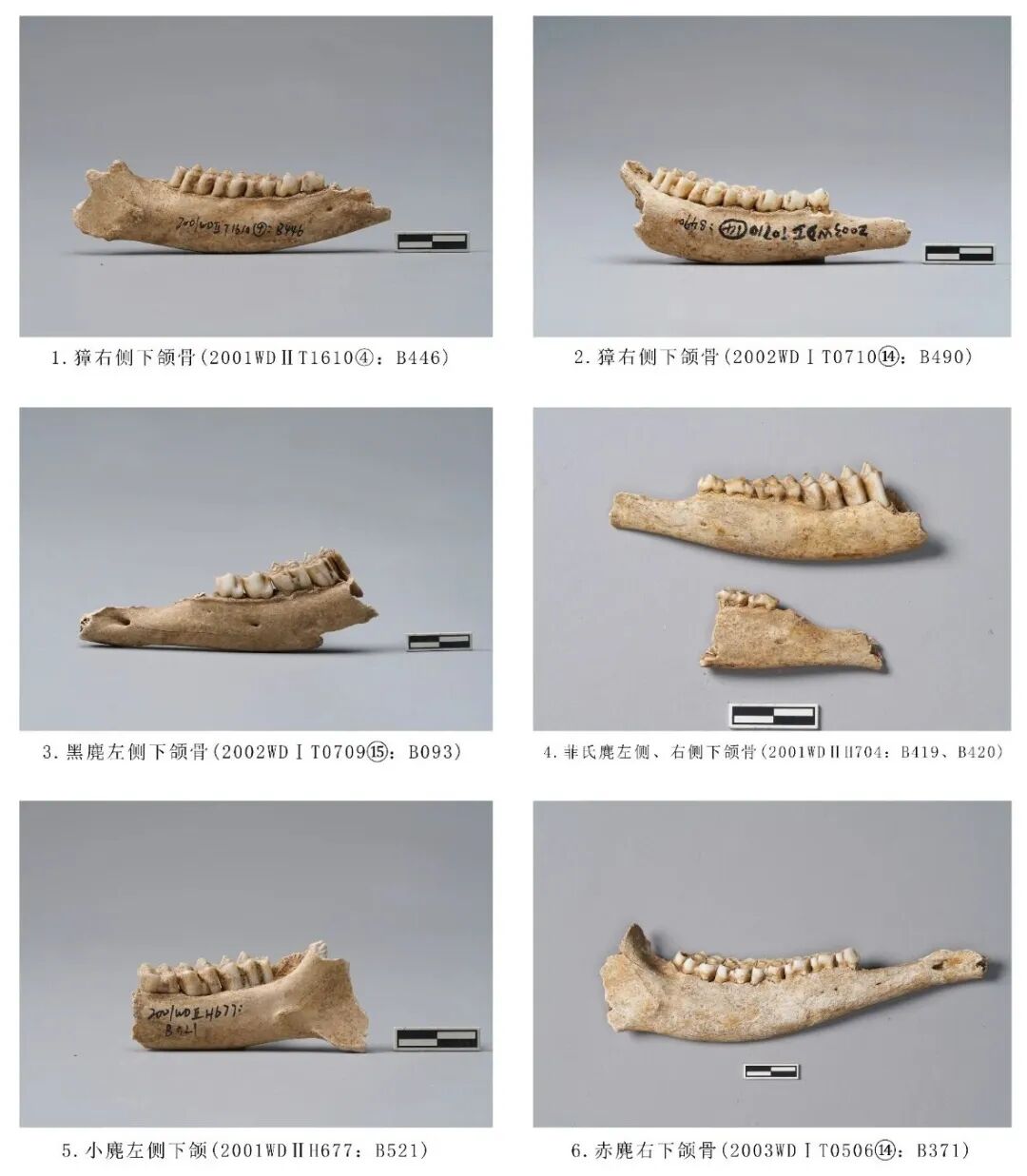
图四 鹿科动物
小麂Muntiacus reevesi
可鉴定标本数122件、最小个体数17。时代包括一至四期、其他新石器时期、商周、东周、汉至六朝、明清时期、次生堆积。骨骼出土部位有游离牙齿、角、下颌骨、肢骨等。
2000WDⅡT1703③:B820,右侧角,次生堆积。角柄部位残。角纤细,角柄部位断面略呈椭圆形,角环突出呈瘤状,角冠向内向后呈弯钩状,纵向布满沟槽未分叉,断面呈椭圆形。角尖端光滑且抛光,为黄色,推测应是使用痕迹。残长90.33、角冠长69.64毫米(图九,5)。
2001WDⅡH677:B521,左侧下颌骨,时代为二期。仅保留水平支后半段和部分角突前半部分,附着M1~M3三枚臼齿。水泥质已完全暴露,且明显内凹低于珐琅质,新月形脊明显。推测其死亡年龄应处于中年阶段。残长59.9、M1~M3长33.53毫米(图四,5)。
菲氏麂Muntiacus feae
可鉴定标本数10件,最小个体数7。时代包括骨骼一至三期。分布部位主要为下颌骨。
2001WDⅡH704:B419,左侧下颌骨,时代为二期。保存大部分水平支,附着P2~M2五枚臼齿,水平支骨骼有轻微的烟熏痕迹,P4有染灰色现象。从P2齿冠咬合面远中面开始,臼齿的磨损呈现出一条内凹的弧线,P4齿冠最低矮,磨损最为严重,M1开始逐渐上升。水泥质完全暴露且内凹,颊侧齿尖的位置明显低于舌侧齿尖,新月形脊明显。推测其死亡年龄应处于中年阶段。残长79.41、臼齿齿列长45.98毫米(图四,4)。
黑麂Muntiacus crinifrons
可鉴定标本数3件、最小个体数为2。时代主要集中在二、三期,骨骼部位主要为下颌骨。
2002 WDⅡT0709⑮:B093,左侧下颌骨,时代为二期。残存水平支前段,P2断裂,保留齿根,附着P3~M2四枚臼齿,M2破损。齿冠所有水泥质都磨损暴露,从磨损痕迹分析该个体的死亡年龄应处于中年阶段。残长99.84、齿列长44.72毫米(图四,3)。
獐Hydropotes inermis
可鉴定标本数6件,最小个体数5。时代包括一至四期。出土骨骼主要为下颌骨。
2001WDⅡT1610:B446,右侧下颌骨,时代为二期。水平支前段缺失,保留部分上升支,附着P3~M3五枚臼齿。骨骼整体纤细,水平支呈弧形,在M1对应处最为低矮,与上升支的联合处略有突起,往角突方向内凹;齿冠均有磨损,水泥质内凹,颊侧齿冠低于舌侧齿冠。从牙齿磨耗程度分析该个体的死亡年龄应处于中年阶段。残长93.68、齿列长45.96毫米(图四,1)。
2003WDⅠT071014:B490,左侧下颌骨,时代为三期。保留水平支和全套臼齿。P2除近中齿尖无磨损之外,所有臼齿均暴露水泥质且内凹,从牙齿磨耗程度分析该个体的死亡年龄应处于中年阶段。残长89.37、齿列长52.96、M1~M3长31.02毫米(图四,2)。
(2)牛科 Bovidae
水牛Bubalus sp.
可鉴定标本数244件,最小个体数14。时代包括一至四期、其他新石器时期、商周、次生堆积。出土骨骼部位主要有游离牙齿、下颌骨、脊椎骨、肢骨等。
2002WDⅠT070826:B094,水牛右角角心,时代为二期。后侧骨骼破碎,从前侧观保存较好。大多数鹿科动物的角心上覆盖一层愈合的毛发,它们的角每年脱落一次,而牛科动物有真正的角,由头骨上额骨的骨质突起(角心)以及上面覆盖发角质鞘构成[5]。表面分布粗糙的条状纹路,远端纹路更为致密,断面呈圆卵型,内部有结构不一的隔膜将角心分隔成多个空腔。全长249.98、近端最大宽120.45 毫米(图五,5)。
2002WDⅡT070720:B041,右侧下颌骨,时代为二期。保存较差,仅保留水平支中段,附着P2~M2五枚臼齿,臼齿有松动现象。齿冠较低,磨损较为严重,珐琅质暴露,牙釉质明显高于珐琅质,M1、M2颊侧齿冠前叶与后叶分别有灰黑色小点,应是与上颌臼齿磨损导致。从牙齿的磨损程度推测,该个体的死亡年龄应为老年阶段。残长166.41、臼齿长114.29毫米(图五,6)。
2000WDⅠT070427:B169,右侧掌/跖骨,时代为一期。骨骼整体保存较为完整,仅在近端关节面和远端滑车上端位置有破损,前端有1条、后端有2条裂纹贯穿整个骨干。长205、近端关节宽63.3、远端关节宽73.8毫米(图五,3)。
2003WDⅠT050824:B642,左侧股骨,时代为二期。保存骨骼近端关节以及部分骨管。大转子明显高出关节面,股骨头有凹陷,转子窝斜行,向下终止于小转子,小转子有明显凸起。骨骺生长线未完全愈合,能辨出其生长方向,推测其死亡年龄处于青年阶段。在近中靠近关节头的骨管部位有两处明显的痕迹,骨管中部有明显的砍砸痕迹,且围绕骨管一周均有砍砸痕,推测其折断骨管的方式应为环绕型砍砸痕迹[6]。残长209.1、近端关节最大宽145.6毫米(图五,4)。
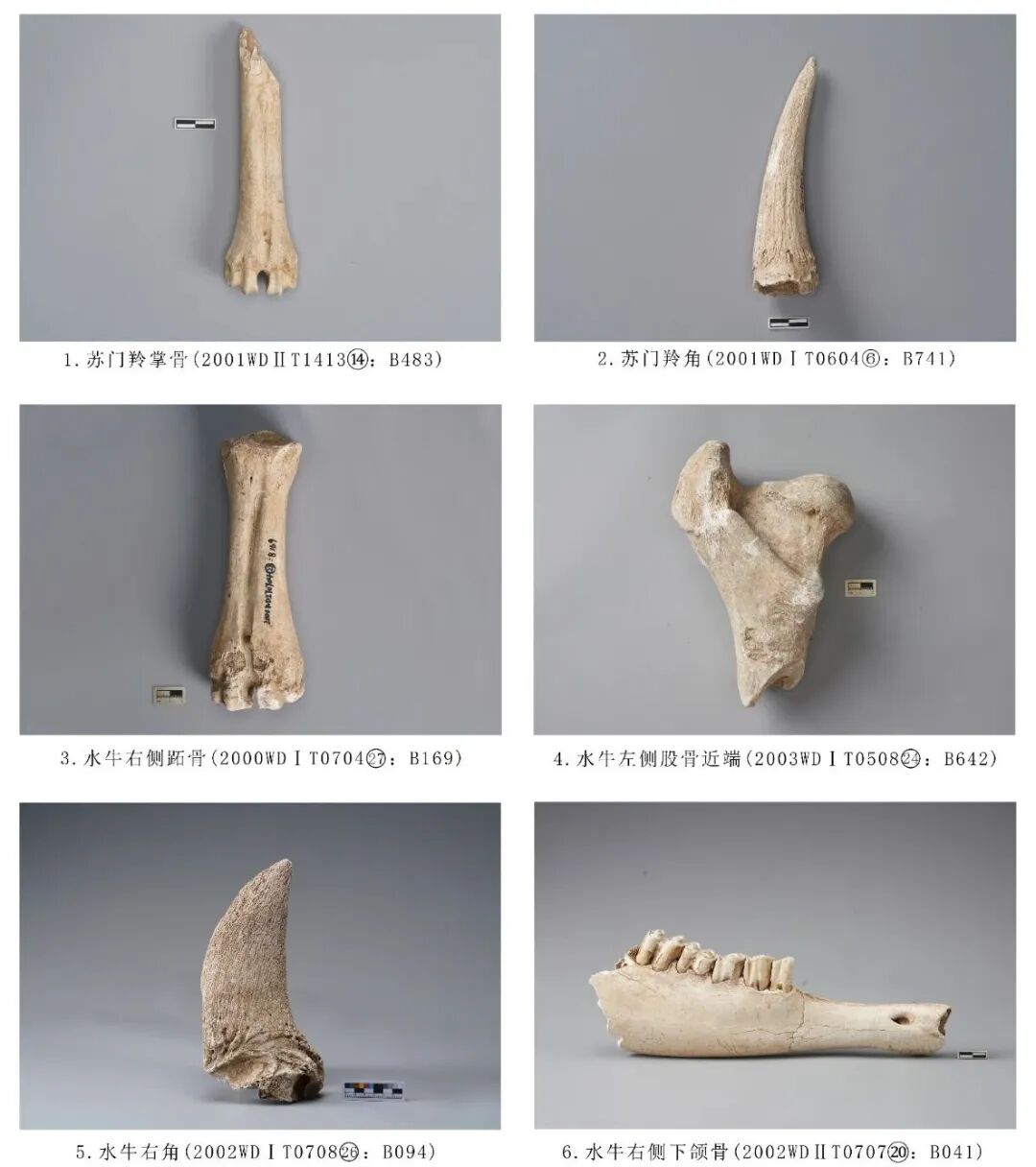
图五 牛科动物
苏门羚Capricornis sumatraensis Bechstein
可鉴定标本数共计12件,最小个体数7。时代包括一至四期、其他新石器时期。出土骨骼主要为角、掌跖骨、桡骨、胫骨等。
2001WDⅠT0604⑥:B741,左角,时代为四期。保存较好。角整体呈锥形,角的基部呈圆形,向上逐渐内缩成尖角,向后上方倾斜,表面布有短小细密的沟槽。长134.77、角基左右径35.34、前后径37.31毫米(图五,2)。
2001WDⅡT1413⑭:B483,左侧掌骨,时代为二期。骨骼近端关节缺失,骨干断面新鲜,应是发掘时所致。骨骼整体宽扁,前侧的血管沟槽宽而深,滋养孔为圆形,远端关节面宽,左右两侧均有凹坑。残长140.85、远端关节宽38.26毫米(图五,1)。
羊亚科Caprinae
未定种属
可鉴定标本数14件,最小个体数为4。时代包括一至三期、唐宋时期。骨骼分布部位有下颌骨、游离牙齿、胫骨、桡骨、掌跖骨等。
2000WDⅠT0803⑰:B159,左侧下颌骨,时代为一期。残,仅保留部分水平支和M2一枚臼齿。臼齿咬合面磨耗较重,水泥质完全暴露,颊侧齿冠明显低于舌侧。下颌骨残长39.09、臼齿:长×宽11.74×7.4毫米(图六,1)。
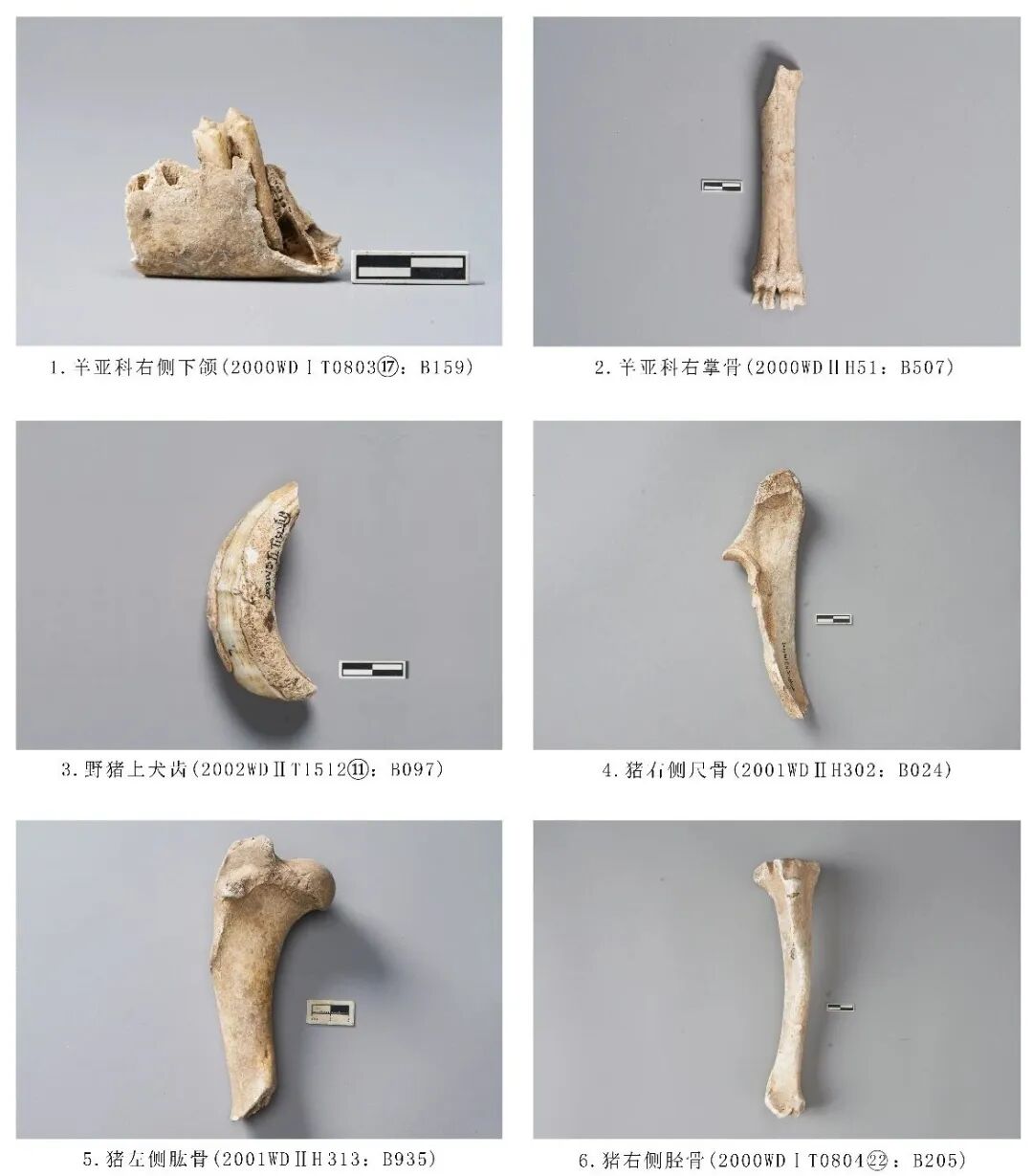
图六 牛科、猪科动物
2000WDⅡH51:B507,右侧掌骨,时代为三期。近端关节外侧残损。骨骼整体形态与苏门羚相似,整体形态更小,前侧血管沟槽不明显,滋养孔呈椭圆形。长132.5、远端最大宽29.21毫米(图六,2)。
(3)猪科 Suidae Gray
家猪Sus scrofa f. domestic
可鉴定标本数319件,最小个体数26。一至四期、其他新石器时期、商周、东周、汉至六朝、东周、明清各个时代均有出土。骨骼分布部位包括游离牙齿、上下颌骨、脊椎骨、肩胛骨、肢骨等部位。
2001WDⅡH302:B023,下颌骨,时代为三期。保留左右下颌骨水平支及前端吻部联合处,左侧附着P2~M3六枚臼齿,右侧附着P4~M3四枚臼齿,保留P2、P3齿槽。水平支较高、平直,臼齿齿列扭曲,M1、M2向外倾斜。牙齿磨耗严重,除M3外,已不见齿冠咬合面珐琅质,M3左侧磨损比右侧更严重,右侧呈片状磨损状。从牙齿磨损程度分析,该个体应处于老年阶段。残长170.73、左侧齿列长105.70,右侧齿列长84.93、M1~M3长72.29、M3长×宽:40.83×17.91毫米(图七,3、4)。
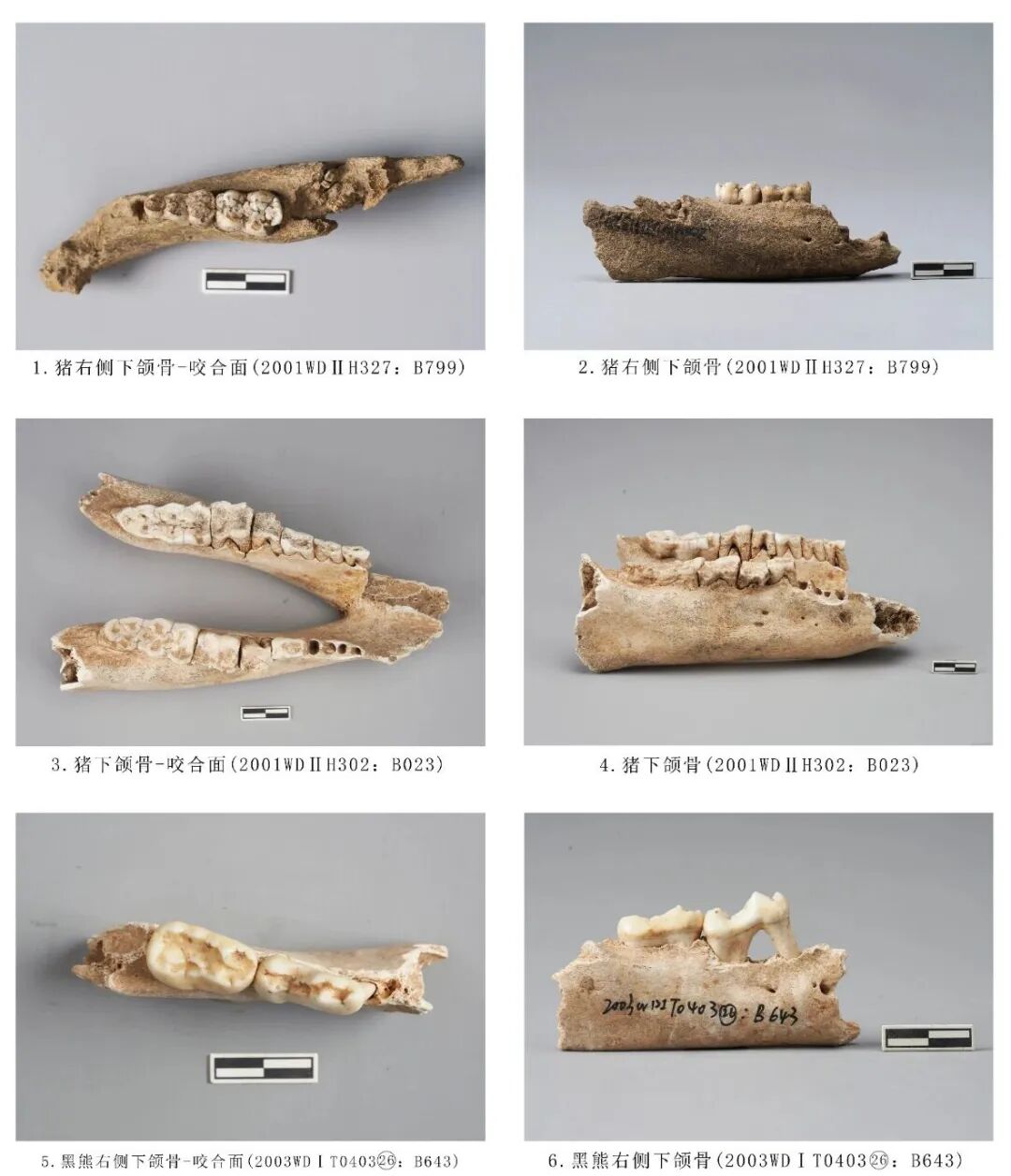
图七 猪科、熊科动物
2001WDⅠH327:B799,右侧下颌骨,时代为三期。保留水平支,附着P4、M1两枚臼齿。骨骼整体呈灰黑色,烟熏痕迹较为严重,臼齿有染黄色现象。下颌骨整体发育较小,臼齿呈点状磨损,从骨骼形态来看,该个体应处于幼年阶段。残长107.93、臼齿齿列长32.96毫米(图七,1、2)。
2000WDⅠT080422:B205,右侧胫骨,时代为一期。保存完整。胫骨脊高耸且弯曲,髌骨系带在胫骨结节处内凹,肌腱沟夹角呈锐角,胫骨粗隆不显著,内髁隆位于尾侧面内侧,内髁显著,远端关节面呈梯形。全长208.24、近端关节宽54.43、远端关节宽30.38毫米(图六,6)。
2001WDⅡH313:B935,左侧肱骨,时代为三期。保留近端关节以及部分骨管。大结节大于、高于小结节,肱骨头圆滑,没有明显的肱骨颈,三角肌粗隆有发育。残长165.58、近端关节最大宽50.14、厚65.36毫米(图六,5)。
2001WDⅡH302:B024,右侧尺骨,时代为三期。远端残。鹰嘴突窄而高,呈半圆形,喙突不明显,骨干侧扁。残长149.75、近端关节宽32.63毫米(图六,4)。
野猪 Sus scrofa
可鉴定标本数11,最小个体数7。时代包括二、三期、商周时期。可鉴定出的动物骨骼主要为犬齿。
2001WDⅡT1512⑪:B097,右侧上犬齿,时代为二期。齿根残。近中面的咬合面宽而长,前侧有带状的齿质条带。牙齿整体呈“C”形,向上、向外倾斜。长71.35、前后径26.74、左右径16.99毫米(图六,3)。
3.食肉目Carnivora
共出土6科共计8种动物,出土骨骼共计1538件,骨骼部位包括游离牙齿、上下颌骨、肢骨等部位。
(1)猫科Felidae
虎Panthera tigris
可鉴定标本数8件,最小个体数为3。时代为二、三期。出土的骨骼部位有犬齿、下颌骨、肱骨、跟骨等。
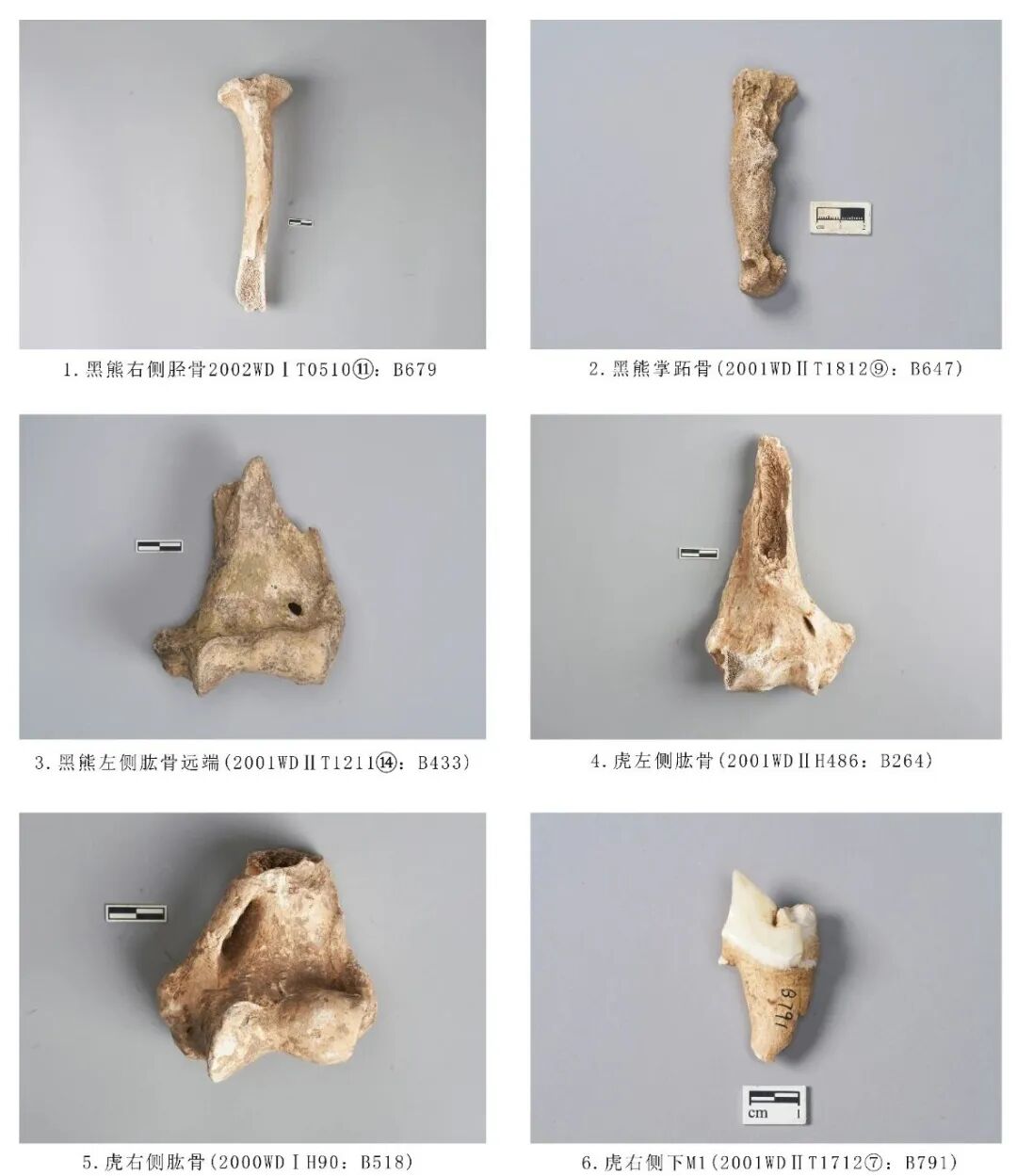
2001WDⅡT1712⑦:B791,右侧下颌M1,时代为三期。仅保留臼齿前段,后段缺失,下前尖崩残。肉食动物裂齿的齿冠沿着两个高齿尖的脊呈明显的刀片状,并维持着一定的高度[7]。残存的齿尖位置未见明显的磨损痕迹,推测该个体的死亡年龄应处于青年阶段。齿冠残长21.21、高40.16毫米(图八,6)。
图八 熊科、猫科动物
2001WDⅠH486:B264,左侧肱骨,时代为二期。保存远端关节部位。整体粗壮且侧扁。远端关节面的滑车较扁,嵴棱隆起不明显,内上髁明显向内伸展,内上髁上方有一髁上孔呈长扁椭圆状,鹰嘴突呈三角形,鹰嘴窝较浅,略呈水滴状。残长142、远端关节最大宽82.5毫米(图八,4)。
2000WDⅠH90:B518,右侧肱骨。残,仅保留远端关节部位。滑车上方有剔肉性痕迹。残长76.68、远端关节最大宽76.68毫米(图八,5)。
豹 Panthera pardus
出土骨骼仅有2件,可鉴定标本数2、最小个体数2。时代为一期和其他新石器时期。骨骼为胫骨和跟骨。
2001WDⅡH173:B404,右侧跟骨,其他新石器时期。骨骼保存完好,外侧和后侧部分有数条较深的砍砸痕迹。近端关节面内凹,载距突未及后缘,前突较短且略平直。全长103.26、近端关节最大宽33.3、远端关节宽25.52毫米(图九,4)。
(2)灵猫科Viverridae
花面狸Paguma larvata Hamilton-Smith
可鉴定标本数2件,最小个体数为2。时代为大溪文化三期、其他新石器时期。骨骼部位主要为下颌骨。
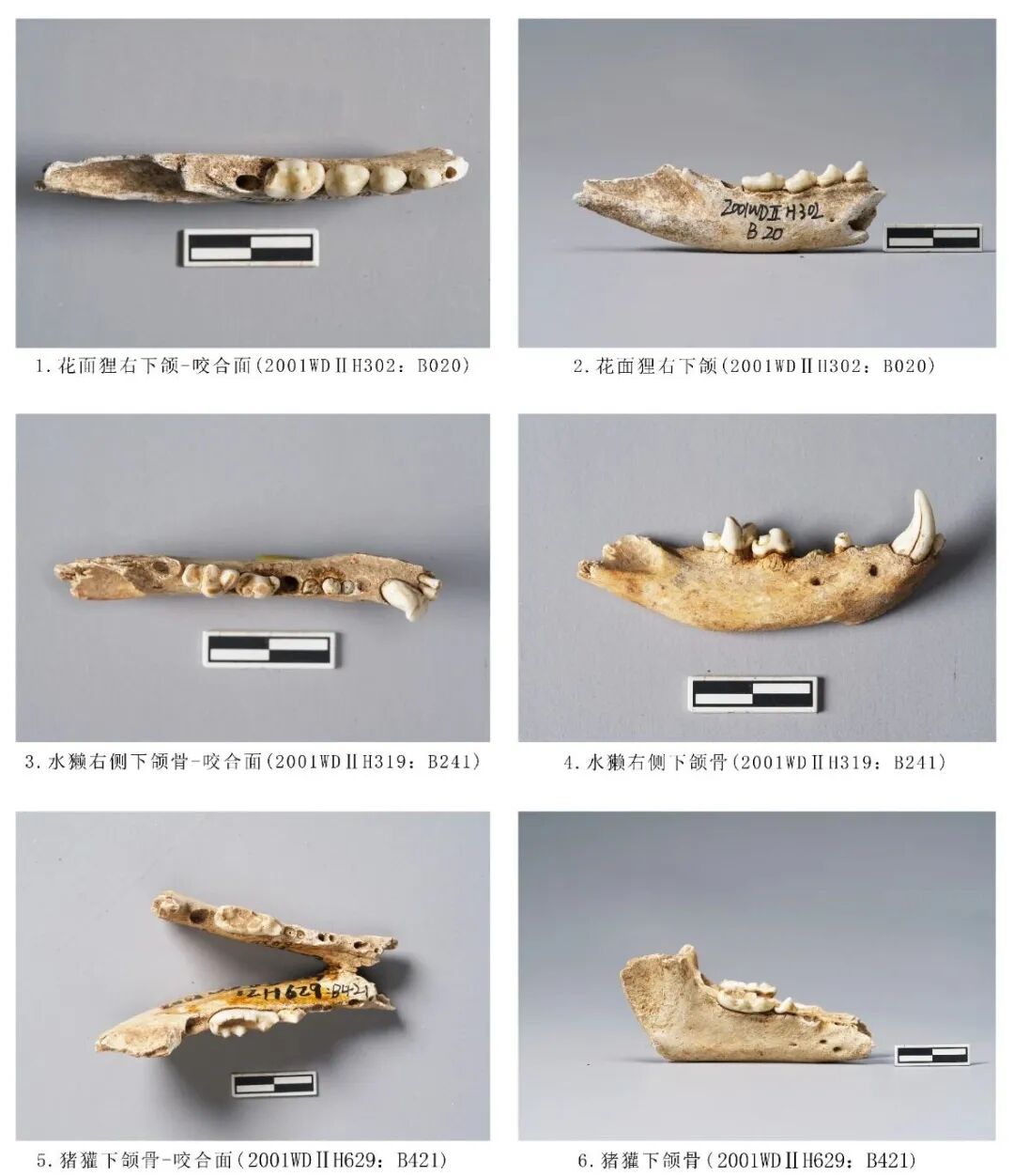
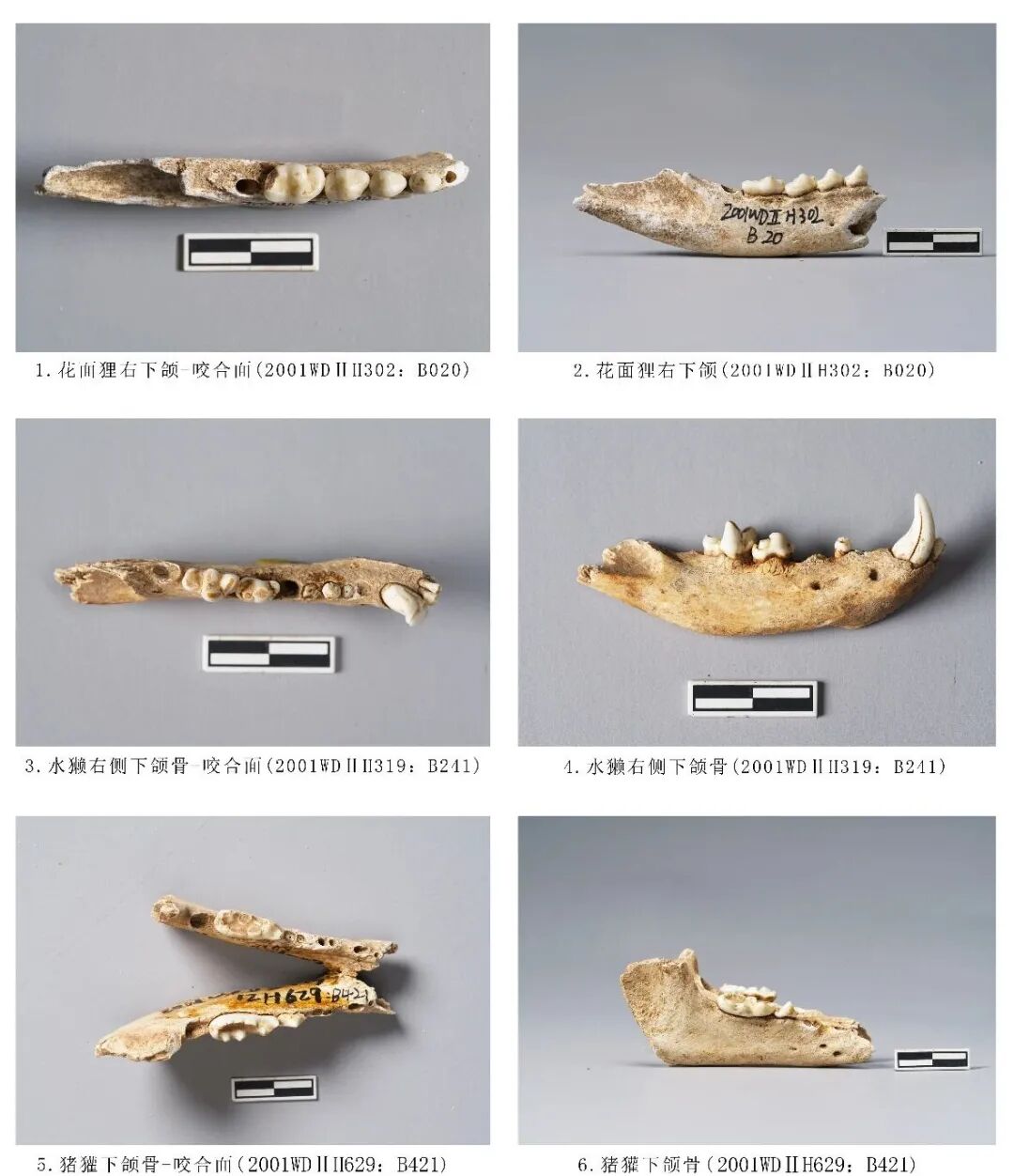
2000WDⅡH108:B510,右侧下颌骨,时代为新石器时期。前端水平支残损,保留大部分水平支和上升支。附着P2、M1、M2三枚臼齿,M2缺失,保留齿槽。P2整体向近中面倾斜,有一个突起的齿尖;M1下原尖大于下前尖和下后尖,各齿尖均低平,下前尖横列呈刃状,下内尖和下次尖不明显;M2颊侧齿冠前端崩残,咬合面磨损严重,呈颊侧高舌侧低的状态,且已不见珐琅质,在舌侧靠后位置的齿根上端形成一个明显的磨损台面,但其他两枚臼齿仅在齿尖位置有轻微的点状磨损痕迹。从牙齿的磨耗程度分析,该个体的死亡年龄应处于中年至老年阶段。残长68.1、齿列长26.5、M1长×宽:6.38×4.83、M2:长×宽9.11×4.15毫米(图九,6)。
2001WDⅡ H302:B020,右侧下颌骨,时代为三期。水平支前端残损,保留大部分水平支和上升支。附着P3~M2四枚臼齿,M3缺失,保留齿槽,M2臼齿远端部分崩残。臼齿磨损较轻,仅M3的下原尖有磨损,说明该个体的死亡年龄应处于青少年阶段。残长51.42、齿列长20.3、M1长×宽9.48×5.18毫米(图一〇,1、2)。
图一〇 灵猫科、鼬科动物
(3)熊科Ursidae
黑熊Ursus thibetanus
可鉴定标本数21件、最小个体数为6,时代为二、三期、其他新石器时期、商周、明清时期。骨骼分布部位有下颌骨、游离牙齿、犬齿、肢骨等。
2003WDⅠT040326:B643,右侧下颌骨,其他新石器时期。仅保留部分水平支,附着P4、M1两枚臼齿,P4对应水平支前后位置分布有2个下颏孔。水平支较平直,臼齿的釉质层较厚,臼齿整体偏窄长。P4暴露2齿根,颊侧的下前尖、下原尖、下次尖,舌侧发育下前附尖、下内尖、下后尖,这6个齿尖相连成脊,围成一个窄而深的内凹面;颊侧3齿尖之间有明显的齿沟,远中齿沟由下次尖与下后尖相连,形成一个宽的三角跟座;未暴露齿质点。M1三尖座略高于跟座,齿尖整体低矮,且齿脊也尖锐;舌侧齿尖高于颊侧齿尖,下原尖、下次尖分布于颊侧,下前附尖、下前尖、下后尖位于舌侧,5齿尖围成的内凹面也有隆起的较为尖锐的齿脊;未暴露齿质点。从牙齿的磨耗程度分析,该个体的死亡年龄应处于青年阶段。残长75.89、水平支高27.37、齿列长41.79、M1长×宽:20.84×12.02毫米(图七,5、6)。
2001 WDⅡT1211⑭:B433,左侧肱骨,时代为商周。骨干部分被砍断,仅保留远端部分,内上髁有缺损。骨骼整体较侧扁,上髁脊发达,内上髁向内伸展成平面,并遮盖住部分鹰嘴窝;滑车矢状脊发育圆钝,不明显,滑车上孔呈椭圆状。残长111.54、远端关节最大宽85.71毫米(图八,3)。
2002WDⅠT0510⑪:B679,右侧胫骨,时代为三期,右侧胫骨。保存近端关节和部分骨管,骨管有裂纹。髁间隆起于近端关节面中间相交,胫骨粗隆微内凹,内侧髁圆平滑,呈半圆形。残长207.02、近端关节宽66.78毫米(图八,1)。
2001WDⅡT1812⑨:B647,掌/跖骨,时代为二期。保存较好,无明显缺失部位。在骨体外侧(左侧)部位有明显不正常的圆状孔和圆窝,并有骨骼突出骨体,这是明显的骨质增生病变,这种骨骼枝稍部位的病变并未对动物造成致命伤害,是动物在自我治疗中逐渐恢复的表现[8]。全长98.48毫米(图八,2)。
大熊猫Ailuropoda melanoleuca
可鉴定标本数2、最小个体数2。时代为一、六期。骨骼部位为上颌骨和尺骨。
2000WDⅠT0503⑪:B152,右侧上颌骨,时代为六期。保存不完整,仅保留上颌M3一枚臼齿以及部分上颌骨,M3舌侧略崩残。大熊猫臼齿最显著特征是齿冠为较复杂的丘形齿,齿冠面齿突、皱纹较多[9]。该枚牙齿整体形态粗壮,颊侧齿冠部分与齿根相比,明显鼓出;舌侧齿冠向内收,形成一条明显的齿带;冠面呈方圆形内凹;近中面边缘平直;远中面边缘较圆滑。磨损痕迹不明显。残长66.7、M2长×宽:21.35×24.45毫米(图九,2)。
图九 鹿科、猫科、灵猫科、熊科动物
2000WDⅠT080423:B195,右侧尺骨,时代为一期。保存近端关节和部分骨干。近端鹰嘴突膨大而向内侧伸展强烈,沟状突强烈向外伸展,半圆切迹浑圆,关节面扭曲,喙突平坦无折面。残长164.58、近端关节最大宽49.32毫米(图九,1)。
(4)犬科Canidae
狗Canis lupus familiaris
可鉴定标本数1110件,最小个体数为32。时代包括大溪文化时期一至三期、其他新石器时期、商周、汉至六朝、唐宋时期。骨骼分布部位有头骨、上下颌骨、游离牙齿、肋骨、肩胛骨、脊椎骨、肢骨、盆骨、掌/跖骨、指/趾骨等。
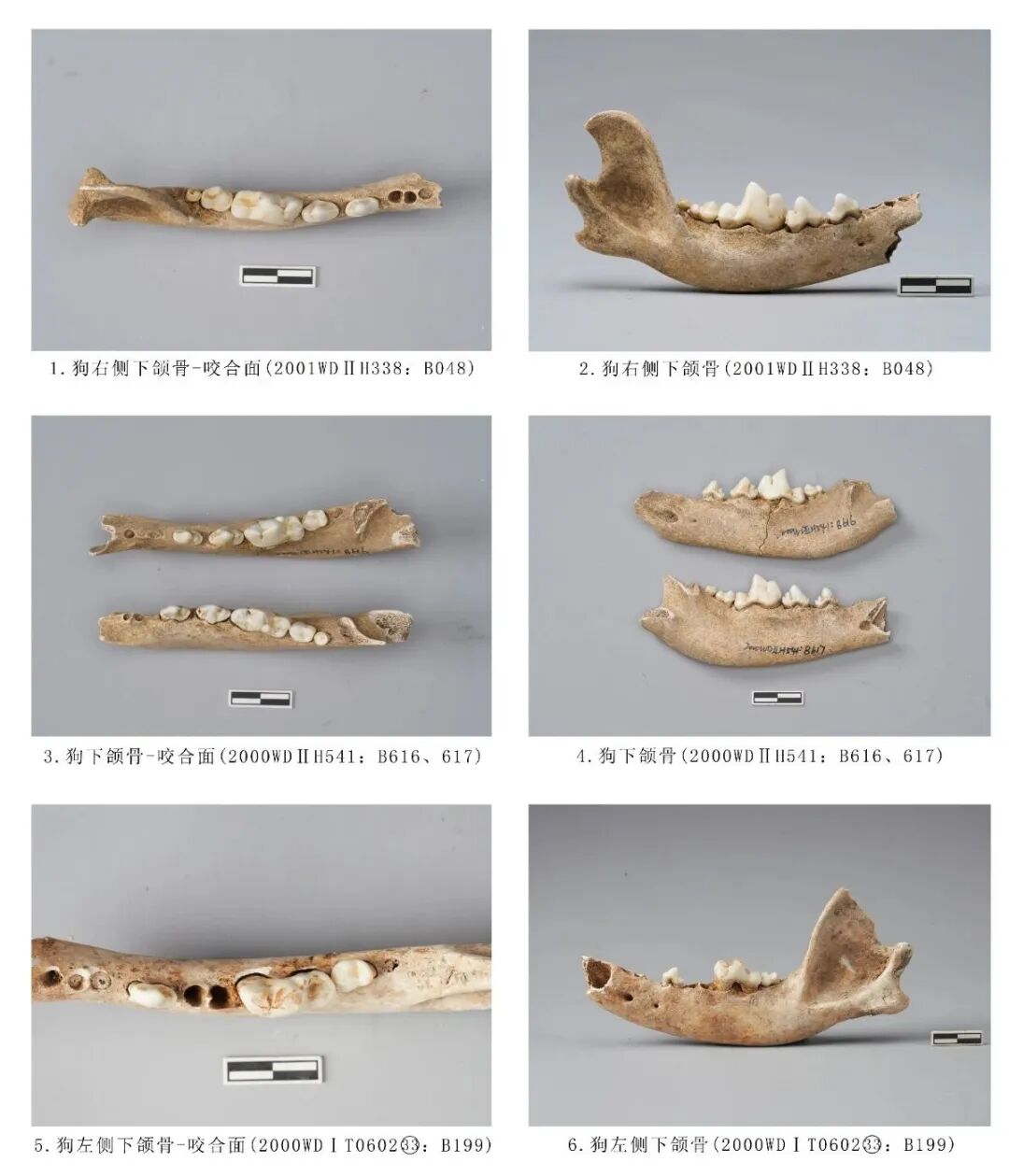
2000WDⅠT060233:B199,左侧下颌骨,时代为二期。保存水平支和上升支,冠状突部分缺损,保留P3、M1和M2三枚臼齿,门齿、犬齿、P1、P2、P4保留齿槽。P3齿尖未出现磨损痕迹;M1前尖与原尖磨损严重,呈颊侧低舌侧高的斜坡状,后尖呈带状磨损,舌侧内尖呈点状磨损,次尖无磨损痕;M2原尖呈点状磨损,其余齿尖均未磨损;M3未萌出。推测死亡年龄应为中年阶段。全长143.92、残高57.29,M1长×宽:20.45×7.65毫米(图一一,5、6)。
图一一 犬科动物
2001WDⅡH338:B048,右侧下颌骨,时代为二期。水平支前段缺失,保留后段水平支和完整上升支,P1、P2缺失,保留齿槽,附着P3~M3五枚臼齿,齿尖未磨损,推测其死亡年龄应为青年个体(图一一,1、2)。
2000WDⅡH541:B616、B617,左右下颌骨,左侧保留P1、P2、M3齿槽,附着P3~M2四枚臼齿;右侧保留P3齿槽,附着P3~M3五枚臼齿,时代为二期。臼齿无磨损痕迹,说明该个体的死亡年龄应处于青年阶段(图一一,3、4)
2001WDⅡH173:B406,左侧尺骨,时代为其他新石器时期。保存较为完整,仅在远端关节处有残缺。鹰嘴突平直,顶端较平,半月切迹圆滑,喙突明显突出,骨干细长且微弯曲。残长164.97、近端关节最大宽19.26毫米(图一二,3)。
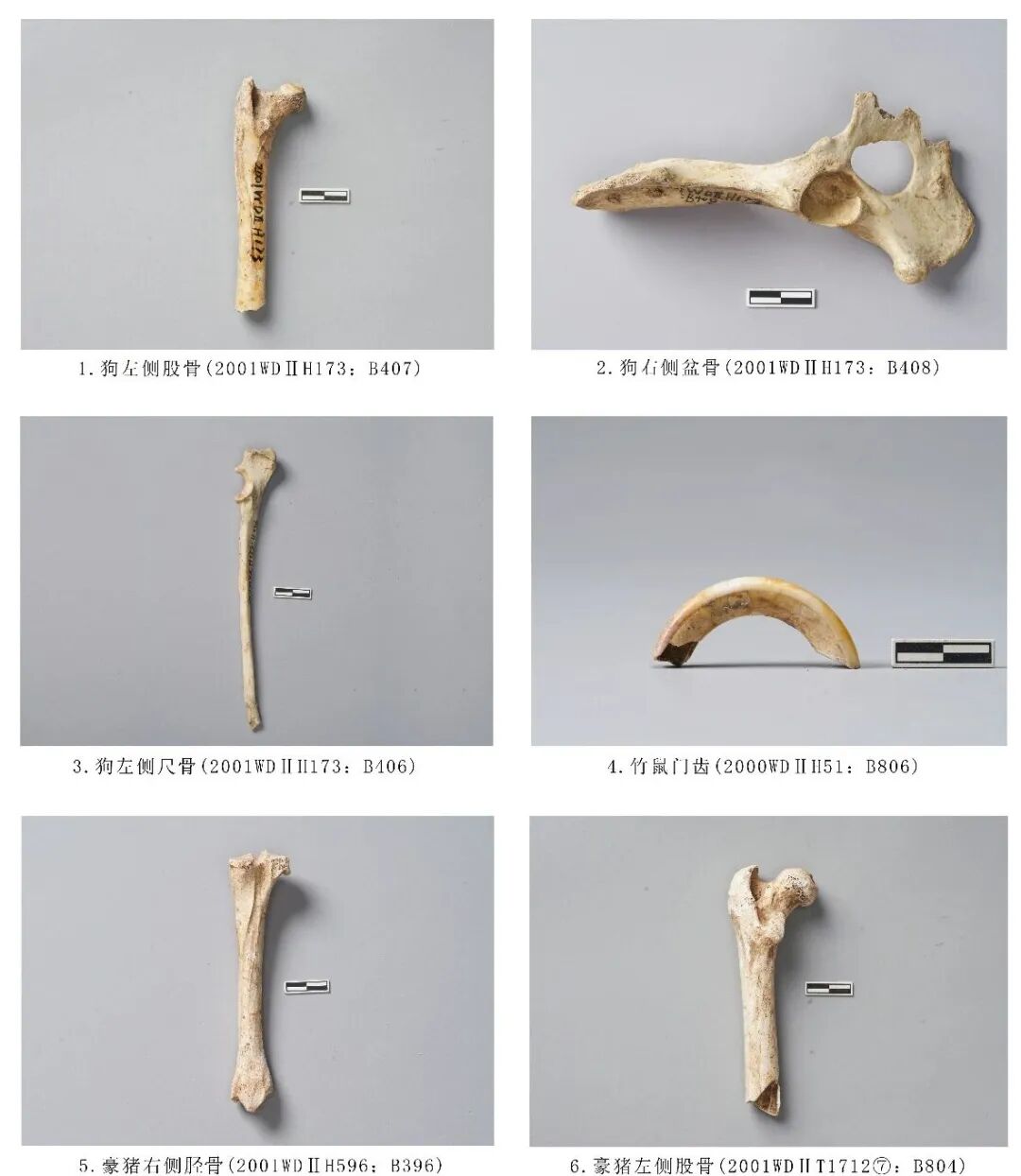
图一二 犬科、豪猪科、竹鼠科动物
2001WDⅡH173:B407,左侧股骨,时代为其他新石器时期。保存较差,仅保留近端关节和部分骨干。大转子与股骨头几乎处于同一水平面,股骨头略高,转子窝平面呈三角形,小转子突起。残长101.73、近端关节最大宽31.03毫米(图一二,1)。
2001WDⅡ173:B408,右侧盆骨,时代为其他新石器时期。保存完整,保留髂骨、髋臼、坐骨、耻骨等部位,闭孔完整,耻骨连接处缺失。残长130.98、坐骨最大宽46.65毫米(图一二,2)。
(5)鼬科 Mustelidae
猪獾Arctonyx collaris
可鉴定标本数148件,最小个体数为33。时代包括一至四期、六期,其他新石器时期、商周。骨骼分布部位游离牙齿、上下颌骨、肢骨等。
2001WDⅡH629:B421,右侧下颌骨,时代为二期。保存完整水平支与部分上升支,角突、髁突等部位均不存。保留犬齿、P1、P2、M2齿槽,附着P3、M1两枚臼齿。猪獾水平支整体较宽、P1与P2之间有齿槽间隙间隙。P3前后排列分别为下前尖、下原尖、下后尖,下前尖与下后尖低矮,下原尖呈刀片状突起;M1的下前尖与下原尖形成三尖座,位置偏颊侧,下原尖大于下前尖,跟座比三尖座低矮,跟座上的5个小齿尖围绕使得跟座中部呈盆形。残长78.78、齿列长24.81、M1长×宽:17.78×7.3毫米(图一〇,5、6)。
水獭Lutra lutra Linnaeus
可鉴定标本数1件,最小个体数为1。时代为三期,出土骨骼为下颌骨。
2001WDⅡ319:B241,右侧下颌骨,时代为三期。保存完整水平支附着2枚门齿,1枚犬齿,M1、M2两枚臼齿,P2~P4缺失,保留齿根。下颌骨整体较纤细、水平支底缘强烈外弧,中间部位最高;残存上升支一小部分咬肌窝;P2前端、P4中部对应的位置分别生长下颏孔。门齿低矮、圆润;犬齿尖锐,两侧有裂纹,无磨损痕迹;M1有3齿尖分布于颊侧:下前尖低矮,下原尖最为粗壮、高大、下附件稍矮于下原尖,3尖分布呈三角形;跟座宽而低矮,上面生长1下后尖;舌侧观下前尖、下后尖分别形成齿沟,下附尖形成一个台面;下原尖与下附尖齿尖磨损最为严重;M2>M1,颊侧分布有下前尖、下原尖和下后尖,3个齿尖均粗壮,下原尖>下前尖>下后尖,3齿尖形成的齿脊将齿冠分为2条宽而深的齿沟,下后尖的远中缘形成一个粗壮的跟座;舌侧下内尖高度与下原尖接近,齿尖整体小于下前尖大于下后尖,齿冠咀嚼面下前尖、下原尖与下内尖呈三角对立;各齿尖均有片状磨损。从牙齿磨耗程度分析,该个体的死亡年龄应处于青年阶段。下颌骨全长62.69、M1、M2长15.47、M1长×宽:7.4×4.49、M2长×宽9.26×5.33毫米(图一〇,3、4)。
4.啮齿目Rodentia
(1)豪猪科Hystricidae
豪猪Hystrix hodgsoni
可鉴定标本数28件,最小个体数为14。时代有一至三期、商周、汉至六朝时期。骨骼部位主要为门齿和肢骨。
2001WDⅡH302:B027,右侧尺骨,时代为三期。远端残。鹰嘴突部分有缺损,整体向内侧倾斜,喙突有一个明显的关节面。残长96.48、鹰嘴突宽15.98、喙突宽17.8毫米(图一五,1)。
2001WDⅡT1712⑦:B804,左侧股骨,时代为三期。远端残。大转子明显高出关节面,股骨头凹陷明显,第三转子突出骨管,骨管平直。残长112.48、近端关节宽40.98毫米(图一二,6)。
2001WDⅡH596:B396,右侧胫骨,时代为商周。保存较为完整,近端关节外侧面有残缺。近端关节面内髁和外髁中间有一条深沟,胫骨粗隆斜向,肌腱沟无夹角,骨管截面呈三角形,内髁不显著。长129.7、近端关节宽31.08、远端关节宽21.94毫米(图一二,5)。
2001WDⅡH596:B409,右侧肱骨,时代为商周。仅保存远端部分骨管和关节部位。骨骼整体较扁圆,滑车较宽,滑车上孔较大,呈椭圆形。残长64.16、远端关节最大宽28.00毫米(图一五,2)。
(2)竹鼠科Rhizomyidae
出土门齿4件,可鉴定标本数4件,最小个体数为3。时代主要为大溪文化时期二至四期。
竹鼠Rhizomys
2000WDⅡH51:B806,门齿,时代为三期。齿根位置破损,齿冠近中面有裂纹。门齿呈“C”型,横截面为内窄外宽的梯形,咬合面不平整。残长42.85、前后宽8.48、左右宽6.4毫米(图一二,4)。
5.灵长目Primates
(1)猴科Cercopithecidae
金丝猴Rhinopithecus roxellanae
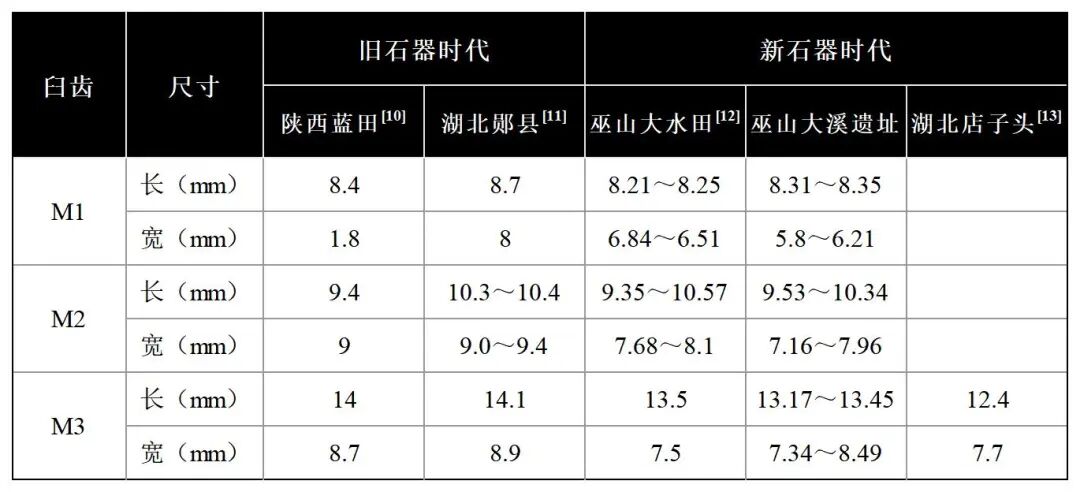
可鉴定标本数13,最小个体数7。时代包括一至四期,其他新石器时期。骨骼部位主要为下颌骨、股骨和游离牙齿(表六)。
表六 金丝猴测量数据对比表
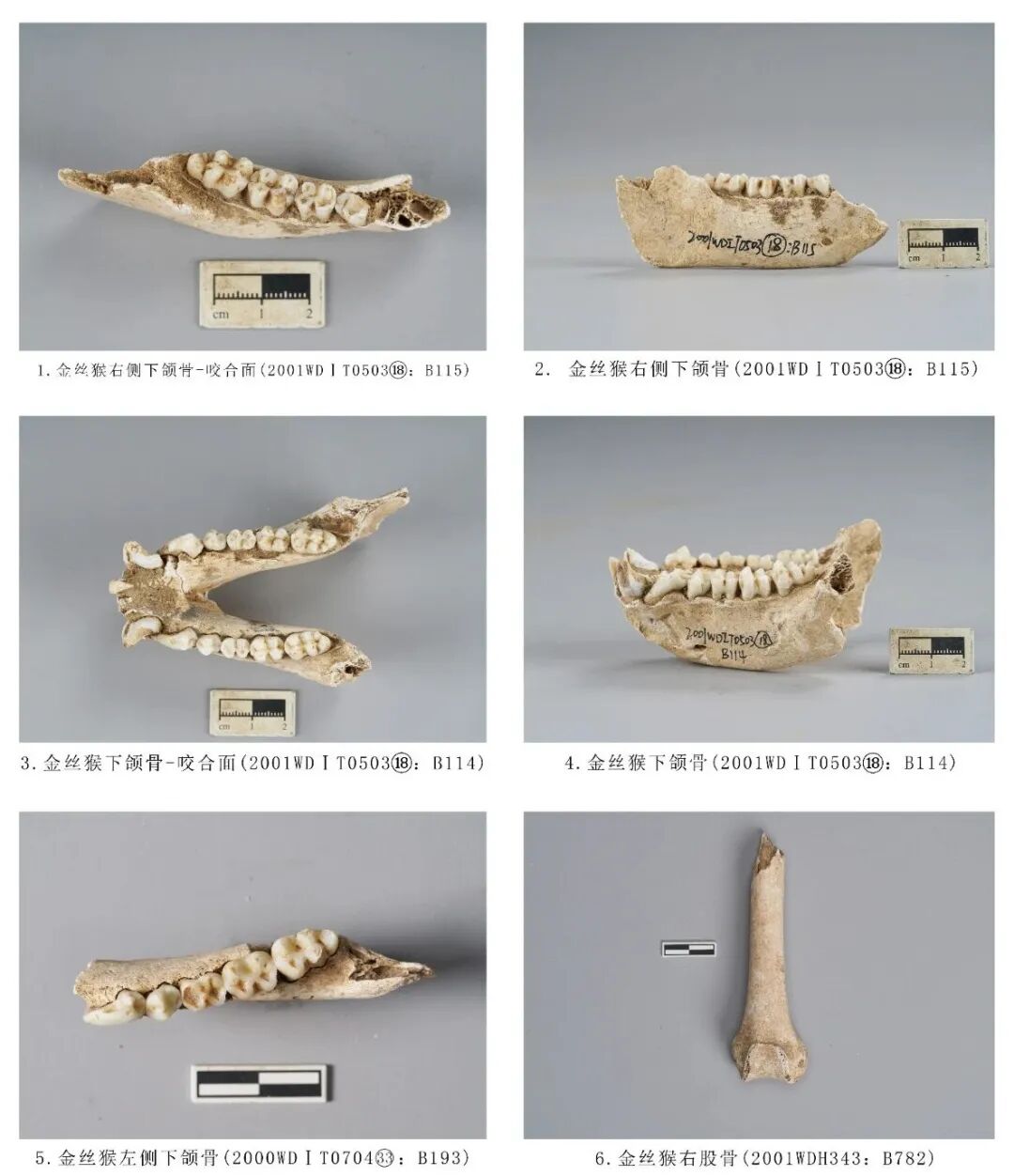
2000WDⅠT050318:B114,下颌骨,时代为三期。保存吻部、水平支部位,有2枚门齿缺失,保留2枚犬齿以及全套臼齿,犬齿有破损。以左侧为例,在P3、M3对应的水平支位置有三道斜直的痕迹,应是发掘所致。下颌骨水平支较高,颊侧观犬齿和前臼齿下方明显向内凹陷;下颏孔较大,位于P3下方。齿式2·1·2·3。门齿呈铲形。犬齿齿冠高而尖锐,齿颈部粗壮且膨出,咀嚼面向颊侧倾斜,近中面有一道较深的凹槽。P3整体向远中方向倾斜发育;颊侧齿尖发育明显高于舌侧齿尖,颊侧下前尖、下原尖(最粗壮)和下后尖之间各有齿脊相连,远中齿脊略长;舌侧的下內尖,略低于下原尖,颊侧三个齿尖分别与舌侧下內尖相连成齿脊,将齿冠的咀嚼面 (明显向舌侧倾斜) 分成三部分,形成三个凹陷面,远中侧凹陷最大,近中侧凹陷次之,中间凹陷最小也最浅,三角形的尖顶均指向下原尖顶端。P4明显臼齿化,发育有颊侧下原尖、下次尖、舌侧下前尖、下后尖4个齿尖,下原尖与下前尖均已出现齿质点,舌侧齿尖的齿质点稍大;下原尖与下前尖高度发达,基本与齿列齿尖平齐,下次尖与下后尖高度较低;下原尖与下前尖分别向前伸出一条齿脊的近中缘,形成小而明显的前凹,四个齿尖形成一个较大的中央凹陷。下臼齿明显地呈双脊型,M1最小,M2居中,M3最大,颊侧齿尖明显低于颊侧齿尖。M1齿尖位置与P4相同,四个齿尖均已出现齿质点,颊侧两个齿尖的齿质点比较大;舌侧齿尖高于颊侧齿尖,咀嚼面向颊侧倾斜;下原尖与下前尖为近中齿尖、下次尖和下后尖为远中齿尖,分别有一条横脊相连,颊侧与舌侧之间的齿尖有较深的齿沟, 将齿冠分成近中和远中两部分;近中缘紧贴齿尖,没有形成前凹,远中缘稍向后伸,形成一个小而明显的后凹。M2咀嚼面呈方形,有4个齿尖,颊侧下原尖、下次尖分别与颊侧下前尖、下后尖在齿冠部分形成两道齿脊,颊侧齿尖与颊侧齿尖分别形成2道宽而深的齿沟;颊侧近中缘与下原尖形成的横脊之间形成齿脊,将前凹形成两部分;4个齿尖围成的最大而深的谷;远中的两个齿尖有明显的跟座,被浅沟分隔而形成颇似两个齿尖的结构;颊侧齿尖暴露齿质点。M3是最大的一枚臼齿,从颊侧观发育有3排齿尖,即颊侧下原尖(略崩残)、下次尖、下后尖,舌侧下前尖、下内尖、下附尖6个齿尖;前边2排齿尖的横脊结构、齿沟与M2相似,远中有一发达的跟座,跟座上发育2个齿尖;未暴露出齿质点,颊侧下次尖与下后尖有磨损痕迹,中间发育有低矮的齿柱[14]。从牙齿磨损的痕程度析,该个体的死亡年龄应处于成年阶段。残长79.3、左侧齿列长49.32、M1~M3长31.7(图一三,3、4)。
图一三 猴科动物
2001WDⅠT050318:B115,右侧下颌骨,时代为三期。吻部及上升支缺失,保存水平支,附着P4~M3四枚臼齿。颊侧观上升支遮住M3远中跟座位置。M1、M2颊侧齿质点暴露比舌侧齿质点更多,M3齿尖有崩残,从牙齿磨耗程度分析,该个体的死亡年龄应处于青年成年阶段。残长77.85、残高30.55、齿列长37.43、M1~M3长30.62毫米(图一三,1、2)。
2000WDⅠT070433:B193,左侧下颌骨,时代为一期。水平支破损,保留P3~M3五枚臼齿,M1~M3有发黄现象,颊侧齿尖均低于舌侧齿尖。P3整体向远中倾斜,倾斜幅度较大,牙釉质向下延伸至水平支颊侧线以下,近中齿根暴露,弯曲呈“C”型,向远中倾斜,颊侧齿尖暴露齿质点;P4颊侧齿尖低于舌侧齿尖,颊侧齿尖暴露齿质点;M1磨损严重,咬合面近中、远中两部分的内凹面水泥质暴露且严重凹陷,是5枚臼齿磨损最为严重的一颗;M2颊侧齿尖2个齿尖的齿质点暴露比舌侧多;M3颊侧下次尖崩残,颊侧的齿尖暴露出齿质点。金丝猴的牙齿,随着年龄的增长,齿冠面逐渐变黄,约4岁性成熟时牙齿开始泛黄(由浅黄至深黄)[15],并结合牙齿的磨耗程度分析,死亡年龄应处于中年阶段,残长55.09、齿列长39.34、M1~M3长26.02、M1长×宽:6.89×5.34、M2长×宽:8.45×6.37、M3:长×宽:10.68×6.63毫米。该个体骨骼形态较小,推测应为雌性(图一三,5)。
2001WDⅡH343:B782,右侧股骨,时代为三期。保存较差,仅保留远端关节和部分骨干。骨骼整体稍扁,滑车位于正中位置,内上髁和外上髁大小结构相似,只是内上髁下方生长一深沟。残长100.02、关节最大宽29.94毫米。残长31.27、关节最大宽33.05毫米(图一三,6)。
金丝猴的伴生动物群,它们均属我国南方广义的大熊猫—剑齿象动物群,其生态环境属于温暖湿润的热带、亚热带环境,但环境气候适应能力较强。
猕猴Macaca mulatta Zimmermann
可鉴定标本数6件,最小个体数为1。出土时代为大溪文化时期二期。骨骼部位分布游离牙齿、下颌骨和肢骨。
2001WDⅡT1814⑫:B431,吻部,时代为二期。仅保存吻部前端位置,附着4枚门齿和2枚犬齿,犬齿齿尖崩残。猕猴体型与金丝猴相比,骨骼形态更为纤细。门齿与犬齿呈左右对称,I1呈铲型,颊侧与舌侧均略呈三角形,舌侧更高,咬合面稍内凹,暴露出齿质点;I2铲形略变形,在舌侧中部位置近中齿冠向远中内收,远中齿冠向外凸出,在中心齿尖的舌侧围成一个小窝;C发育粗壮,为高而尖锐的齿冠,齿颈部膨出,以右侧为例,近中面有不明显的齿脊,齿冠在近中/舌侧上沿齿冠长度方向形成凹沟,齿冠向后倾斜。门齿残长28.42、犬齿齿冠高10.67、门齿齿列长13.04毫米(图一四,4)。
2001WDⅡH481:B613,右侧下颌骨,时代为二期。保留吻部右侧前端、水平支,附着全套右侧下颌牙齿7枚(1枚门齿缺失)。齿式2·1·2·3。下颌骨整体较平直,水平支与吻部交接处平直、圆钝,吻部朝近中延伸,下颏孔位于P3对应位置。门齿呈铲形,稍侧扁。犬齿尖锐,在舌侧靠近远中位置有一台面,应为与上颌犬齿交接处,咬合面有磨损,未至珐琅质。P3颊侧齿冠靠近齿根部位向下倾斜,舌侧有明显的齿带形成跟座,为单齿尖偏向远中方向,齿尖呈三角形。P4在颊侧和舌侧分别分布有下原尖和下前尖,2齿尖大小相似,形成一个短的齿脊相连,下后尖最为低矮,与2齿尖相连成脊连成一个低矮的跟座,倾斜面内凹形成齿冠咬合面。猕猴属上臼齿舌侧、下臼齿颊侧齿尖之间的中裂(中沟)比较浅,牙尖起伏不太大,臼齿形态基本是前宽后窄,前尖与原尖连接的前脊强于后尖和次尖连接的后脊[16]。M1~M3呈臼齿状齿冠面略呈长方形。M1齿冠面有崩残,有5齿尖,颊侧分布下原尖、下附尖、下次尖,舌侧分布下前尖、下后尖。下前尖与下原尖并排,近中缘有齿带;5个齿尖相连成脊,使得冠面形成一个内凹咬合面;5个齿尖均暴露齿质点。M2齿冠齿尖整体较圆钝,颊侧齿尖明显低于舌侧齿尖。颊侧的下原尖、下次尖、舌侧的下前尖(最发育)、下后尖分别形成两条横脊,将齿冠面围成一个内凹面;近中缘高于远中缘;远中齿尖暴露齿质点。M3与M2大小结构相似。从牙齿磨损程度分析,该个体的死亡年龄应处于成年阶段。全长57.81、门齿宽6.53、犬齿齿冠高12.15,臼齿齿列长34.54、M1~M3长22.66、M1长×宽:7.08×5.85、M2长×宽:7.79×6.88、M3长×宽:7.96×7.19毫米(图一四,5、6)。
2001WDⅡT1814⑫:B848,右侧下颌M2,时代为二期。保留2个齿根,近中齿根向内弯曲,远中齿根平直。从牙齿磨耗程度分析,该个体的死亡年龄应处于成年阶段。臼齿长×宽:7.53×6.28毫米(图一四,3)。
山魈 Mandrillus sphinx
可鉴定标本数6,最小个体数2。时代为大溪文化时期二至四期。出土的骨骼均为犬齿(表七)。
表七 山魈犬齿测量数据一览表
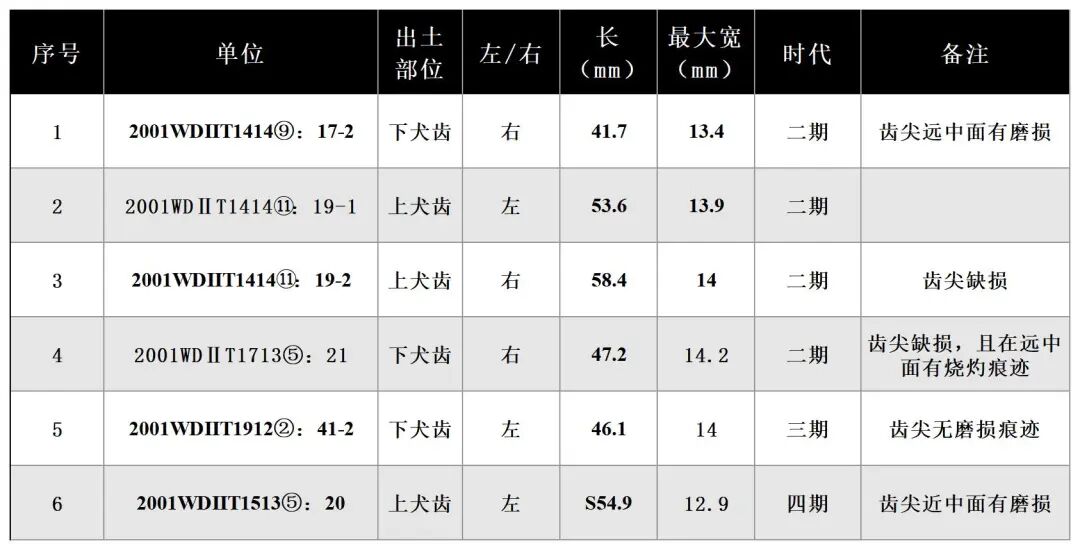
2001WDⅡT1414⑪:19-2,左侧下犬齿,时代为二期。保存完整。牙齿呈弧线型,较为锋利,牙釉质高度与齿根相差不大。齿冠整体内收,断面呈锐利的圆锥形,近中面有一道宽而深的凹槽,远中面有明显的齿沟,后侧有与下颌犬齿摩擦留下的痕迹,咬合面呈锋利的刀片状,近中齿冠有发黄发绿现象(图一四,1)。
2000WDⅡT1503⑤:20,左侧上犬齿,时代为四期。保存完整,近中咬合面有磨损,暴露出牙釉质(图一四,2)。
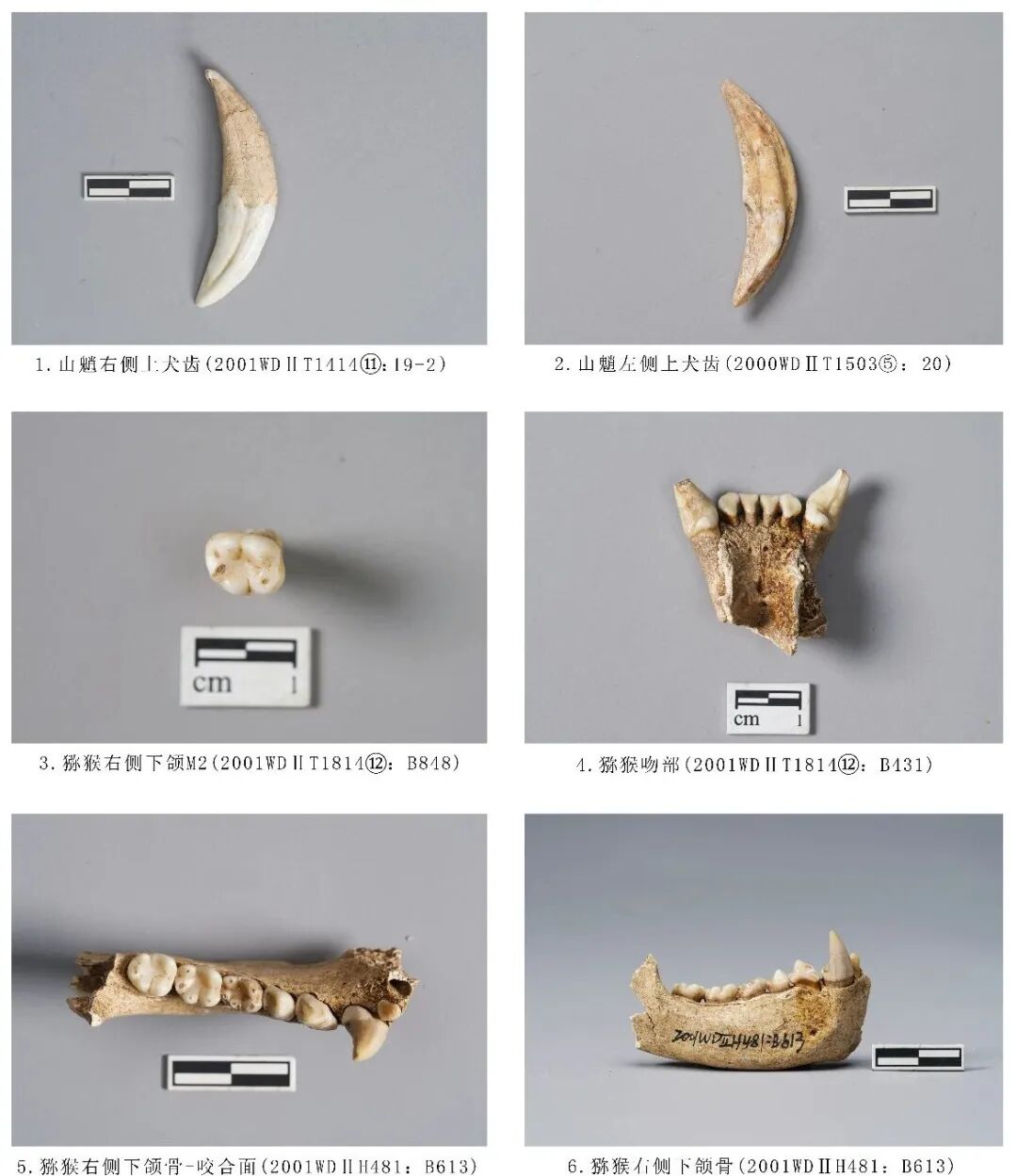
图一四 猴科动物
(二)硬骨鱼纲Osteichthyes
鱼类骨骼在大溪遗址出土的数量共计12441件,占出土动物骨骼总量的51%,可鉴定标本数为542,占鱼类骨骼总数的4.4%。出土的种类有草鱼、青鱼、鱤鱼等,骨骼分布部位及数量主要有下咽骨194件、下咽齿263件。需要说明的是,由于鱼类骨骼的结构系统相当复杂,许多骨骼部位极难保存,并且鉴定的特征性不显著,目前只能对目前只能对下咽齿、下咽骨和特征明显的胸鳍棘等部位进行种属的鉴定,其余部位尤其是椎骨只作数量统计,无法作最小个体数的统计。
1.鲤形目Cypriniformes
(1)鲤科Cyprinidae
青鱼Mylopharyngodon Piceus
出土青鱼下咽193骨件、下咽齿169件,游离牙齿11枚。时代包括一至四期、六期、其他新石器时期、商周、汉至六朝、明清时期。
2000WDⅡH85:B002,右侧下咽骨,时代为三期。保存较为完整,保留3枚下咽齿。整体粗壮,前臂比后壁长,呈三角形微弯曲;内侧较厚,外侧斜出,逐渐变薄;背面有数个大小形状不一的孔洞;咽齿呈臼齿状,前2枚咽齿磨损较轻,第3枚咽齿残,磨损呈内凹状。长94.42、宽69.7、齿列长43.01毫米(图一六,3)。
2003WDⅠT0409⑮:B853,下咽骨,时代为三期。仅保留部分下咽骨并附着2枚臼齿。整体较粗壮,下咽齿粗大而短,有多个隔膜将下咽骨下方分为多个空腔。齿冠呈臼齿状,一大一小,为不规则椭圆形,牙齿釉面光滑,没有槽纹,齿冠面中部有磨损形成的凹坑。残长49.24、齿列长21.45、臼齿长×宽分别为:8.81×12.42、12.64×16.57毫米(图一六,4)。
草鱼Ctenopharyngodon Idella
出土草鱼下咽齿共计54枚,时代包括大溪文化时期二、三期。
2000WDⅡT1303⑬:B850,时代为二期。草鱼下咽齿具有显著的鉴定特征:形态高大侧扁,冠面呈梳形,齿冠顶面倾斜,齿冠中部有一条沟槽,侧面有排列整齐的槽纹。草鱼属只有草鱼1种,是我国特有的鱼类,分布广泛。残高24.58、齿冠最大宽11.28毫米(图一七,1)。
鱤鱼Elopichthys bambusa
出土鱤鱼下咽齿共计18件,时代包括大溪文化时期二、三期。
2000 WDⅡH40:B854,下咽齿,时代为三期。附着一小段下咽骨。下咽齿齿根部位较为粗壮,釉面位于牙齿中部,齿尖收为尖而扁的形状,咀嚼面于齿冠侧面内凹,顶端弯曲为钩状。高26.35毫米(图一六,6)。
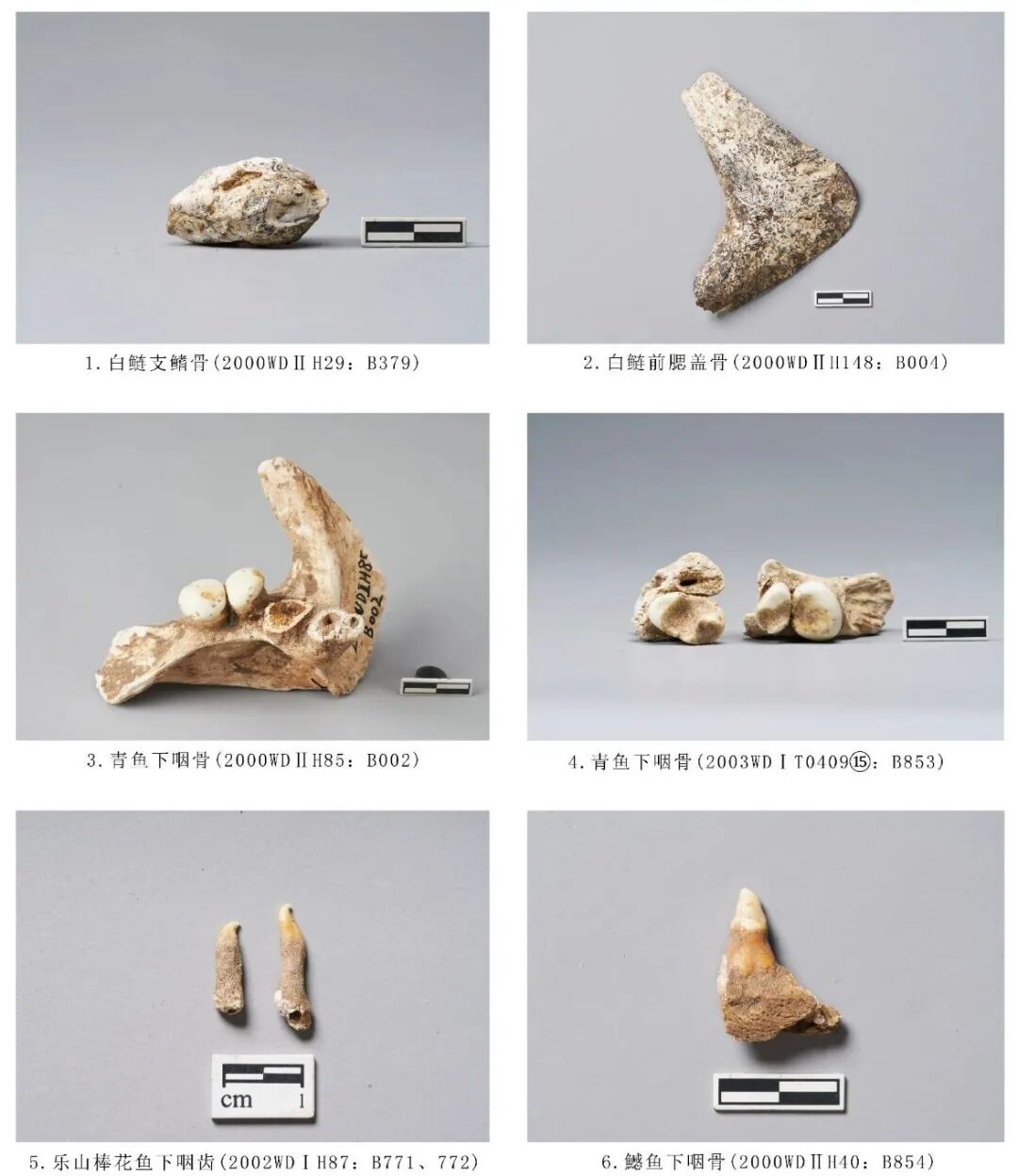
图一六 鲤科动物骨骼
白鲢Hypophthalmichthys molitrix
可鉴定标本数93件,骨骼部位包括下咽齿、齿骨、鳃盖骨、支鳍骨等。时代包括一至四期、新时代时期、汉至六朝、明清时期。
2001WDⅡT1513④:B137,右侧齿骨,时代为四期。背侧与尾侧残。骨骼整体呈三角形,外侧下缘分布有一排圆孔;内侧有一道深沟,深沟下排列分布三个椭圆形小孔。齿骨是位于下颚前端的一对长骨片,前端微向内弯曲,左右齿骨前端相接[17]。
2000WDⅡH148:B004,前鳃盖骨,时代为二期。保存较为完整,两端略残。骨骼呈钝角“V”形,外侧下方分布许多椭圆形小孔。骨骼颜色呈灰黑色,明显为烟熏后的痕迹。残长94.54、残高72.52、宽49.26毫米(图一六,2)。
2000WDⅡH29:B379,支鳍骨,时代为二期。残。呈枣核(橄榄核)状,浑身布满烧灼碳化后的黑色斑点。残长34.49、最大宽20.81毫米(图一六,1)。
白鲢背鳍和臀鳍的支鳍骨中第一支鳍骨形体最大,为一四棱状骨骼,背面的棱(嵴)高而长,腹面的棱低矮而短,一般在白鲢生长的第三年开始膨胀即变异,并随着年龄的增加形态逐渐变化成橄榄核状,并只存在于4龄以上的白鲢[18]。
乐山棒花鱼Abbottina kiatingensis (Wu)
出土下咽齿2件,可鉴定标本数为2。时代为大溪文化时期三期。
2002WDⅠH87:B771,下咽齿,时代为三期。牙齿整体较细长,尖端呈钩状,尖钩锐利。长16.42、宽3.35毫米。2002WDⅠH87:B772,时代为三期。比另一枚牙齿低矮、粗壮,尖端呈钩状,尖钩更倾斜。长11.98、宽3.37毫米(图一六,5)。
2.鲇形目 Siluriformes
(1)鲇科 Siluridae
出土胸鳍棘2件,时代为大溪文化时期三期、唐宋时期。
鲇鱼Silurus asotus,Linnaeus,1758
2000WDⅠT0703③:B816,胸鳍棘,时代为三期。远端残。骨骼整体粗壮,布满密集的纹路。关节面呈半圆形,骨干内侧有一道凹槽贯穿始终,腹侧部分生长有密而突出的尖刺。残长54.36、关节面最大宽20.06毫米(图一五,6)。
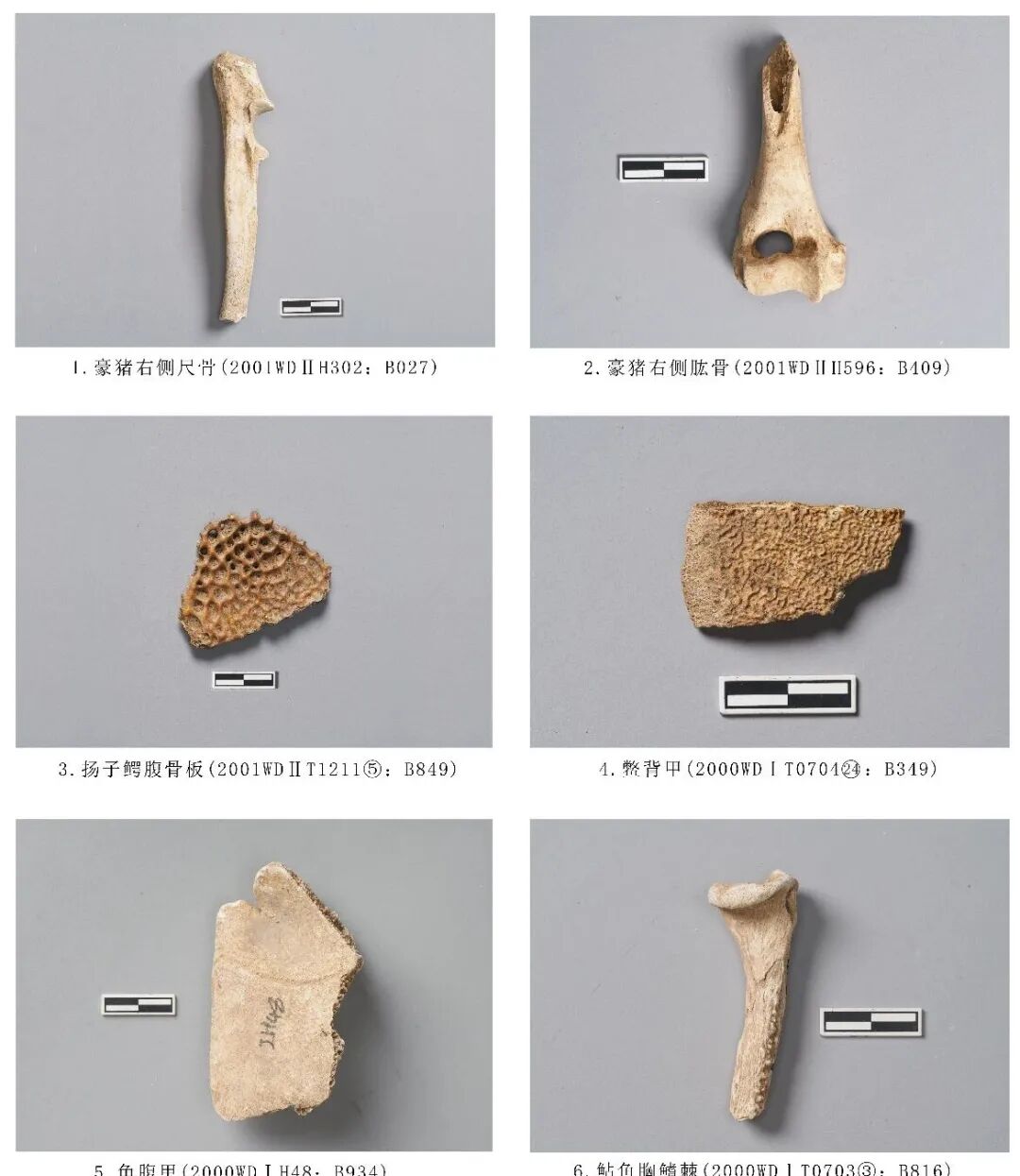
图一五 豪猪科、鲇科、鼍科、鳖科、龟科动物骨骼
(三)鸟纲Aves
遗址内出土的鸟类骨骼数量较少,共计6件,出土的骨骼主要为肱骨、股骨。
(四)爬行纲Reptilia
遗址内出土的爬行类动物的骨骼数量共计263件。出土的动物种类只有扬子鳄、鳖、乌龟3种。
1.鳄目
(1)鼍科Alligatorida
扬子鳄Alligator sinensis Fauvel,1879
出土骨板106件。除东周时期和次生堆积没有出土外,其余时代均有骨骼出土。
2001 WDⅡT1211⑤:B849,腹骨板,时代为商周。整体呈三角形,一条边平直规整,无明显断茬,应是骨板交接处,另外两条边断口明显,骨板整体向内凹。表面布满不规则圆形小坑。长56.71、宽42.29毫米(图一五,3)。
骨板即剥下的鳄皮,鳞片脱落后,骨板仍嵌在皮肤上。背、腹两面的皮肤上均有骨板,背面的骨板方形,有纵列的中棱脊,骨板的上表面有许多形状不规则的凹坑,下表面光滑,微向内凹。腹面的骨板,形状不太规整,没有纵列的中棱脊,表面有许多圆形小凹坑[19]。
2.龟鳖目
(1)鳖科Trionychiade
鳖Pelodiscus sinensis
可鉴定标本数7件,最小个体数为1。均为背甲。时代为大溪文化时期一、二期,商周时期。
2000WDⅠT070424:B349,背甲,时代为二期。残。骨骼较薄,呈弧形,断面一侧平整,应是自然形成,一侧茬口新鲜,应是发掘所致。鳖肋板的宽度大体相等,背面有密集的凹坑,内侧面有一条凸棱。残长34.67、17.4毫米(图一五,4)。
(2)龟科 Mauremys reevesii
乌龟Chinemys reevesii
可鉴定标本数149件,包含背甲、肩胛骨、喙突、肢骨等部位骨骼。时代包括一至六期、汉至六朝、次生堆积。
2000WDⅠH48:B934,腹甲,时代为三期。残。骨骼整体较厚,中间厚、边缘薄,并且两边均分布有弧形的条纹,边缘平整、光滑,为乌龟腹部边缘的骨骼。残长75.66毫米、残宽46.04毫米(图一五,5)。
(五)瓣鳃纲Lamellibranchia
遗址出土瓣鳃纲的总量为30件,共出土3种,均为蚌目蚌科。
1.蚌目 unionida
(1)蚌科Unionidae Rafinesque,1820
圆顶珠蚌Nodularia douglasiae Griffth & Pidgeon, 1833
可鉴定标本数16件,最小个体数为10。时代为大溪文化时期一至四期。
2000WDⅠT070428:B111,右壳,时代为一期。保存较为完整,在后端靠近外侧有部分残缺。贝壳整体中等偏小、窄长,后端圆润,壳质地坚硬,外侧壳顶部位的外皮有脱落,生长线纹路清晰。前端靠近外侧的位置有一完整的圆形穿孔,后端靠近外侧部分因缺失,仅存半圆穿孔,从打孔痕迹看应为双向打孔,即分别从壳的外侧和内侧分别使用工具向中间打孔。圆顶珠蚌栖息环境广泛,在考古遗址中发现频率较高,为优势种[20]。壳长65.54、高11.52、宽34.7、厚4.34毫米(图一七,2)。
剑状矛蚌 Lanceolaria gladiola Heude,1877
2000WDⅠT080326:B935,右壳,时代为一期。保存较差,蚌壳中间断开,顶部、壳边缘有破损。壳质厚、坚固;外轮廓窄长,呈剑状;壳面有规整且清晰的生长纹线。残长186.95、最大宽50.88毫米(图一七,4)。
2000WDⅠT050321:B933,右壳,保存完整,远端有磨损痕迹。整体细小,壳表面凹凸不平,生长纹路清晰。全长89.44、最大宽22.02毫米(图一七,6)。
背瘤丽蚌lamprotula leaii
可鉴定标本数为1,最小个体数为1。时代为大溪文化时期三期。
2001WDⅠT0704⑦:B429,右壳,时代为三期。仅保留壳前端部分,后部残。壳质坚硬,整体较厚,从残存的右壳复原整体应略呈椭圆形。背缘平直,生长纹路明显,壳面除前缘部布满瘤状结节。残长50.97、残宽52.94、壳厚9.54毫米(图一七,5)。
(六)腹足纲 Gastropoda Cuvier, 1797
1.柄眼目Stylommatophora
(1)巴蜗牛科 Bradybaenidae
大脐椰雕蜗牛Coccoglypta pinchoiana (Heude)
可鉴定标本数为1,最小个体数数为1。时代为大溪文化时期二期。
2000WDⅠT0702⑧:B430,蜗牛壳,时代为二期。外层壳破损缺失,残存3层螺层。顶层螺层增长缓慢,随外层逐渐膨大,缝合线深,螺壳布满稠密且细致的生长线。残宽16.67、残高12.73 毫米(图一七,3)。

图一七 硬骨鱼纲及瓣鳃纲
四、 动物种群分析
大溪遗址的时代跨度很大,从大溪文化时期一直延续至明清时期。出土的动物骨骼资料十分丰富,动物种类多样,囊括了哺乳动物、鱼类、鸟类、爬行类、软体动物类5大种类,几乎涵盖全了内陆所有的动物种类。说明五、六千年以来,大溪遗址周围的野生动物资源丰富,淡水资源也很充足,充分体现了该遗址周遭生物的多样性,为大溪遗址先民提供了良好的生存环境。同时,大溪先民也具备一定的家畜饲养能力,农业经济也有所发展。
1.野生动物
遗址时代跨度虽大,但出土的动物骨骼主要还是集中在新石器时代,尤其集中在大溪文化一至三期。这一时期的动物种类变化不大。出土种类数量最多的动物种类为野生生物,哺乳动物共计19种,其中尤以鹿科动物种类最多;鱼类6种;爬行动物3种;软体动物4种。该遗址的动物种群大多与晚更新世的“大熊猫——剑齿象动物群”相同。大熊猫——剑齿象动物群包括竹鼠、华南豪猪、猪獾、大熊猫巴氏亚种、最后鬣狗、爪哇豺、黑熊、华南虎、东方剑齿象、华南巨貘、中国犀、野猪、麂、獐、水鹿、青羊、苏门羚、水牛等,而遗址内除了最后鬣狗、爪哇豺、东方剑齿象等动物未见之外,其它的动物种类均在该动物群之列。与此同时,遗址内还有花面狸、水獭、豹、金丝猴、猕猴等全新世的物种,说明大溪遗址在新石器时代中期气候处于过渡阶段,与更新世高温气候较为相似。
史前时期,由于人类对自然的改造能力较弱,动物群体对自然环境有着很清楚的指示意义。对这些动物种类的生活习性稍加分析就会发现不同物种有着不同的生活环境,它们所依赖的自然环境也有所不同。比如鹿科动物栖息的环境主要为阔叶林、针阔混交林、林缘草地及山丘草丛;黑熊和虎为典型的林栖动物,栖息于阔叶林和针阔混交林;犀牛、金丝猴、猕猴等这类动物生活在更为温暖湿润的地区;猪獾生活在阔叶林、针阔混交林的天然岩石裂缝和树洞;竹鼠栖息于竹林、马尾松林及山地阳坡草丛下,为地下穴居动物;豪猪分布很广,为洞穴夜行性动物[21];水獭为半水栖兽类,经常活动于河流、湖泊或溪水中及岸边,为穴居动物;花面狸生活在热带或亚热带山林里,居住于树洞或岩洞中[22]。从这些动物生活的环境来看,三峡地区在新石器时代的植被环境为海拔较高的针阔混交林、海拔较低的阔叶林、灌木丛、草丛,有的动物生活于林间,有的动物栖息于草丛,有的动物生活于竹林。
遗址内发现多种濒临灭绝的珍稀动物,苏门犀、梅氏犀、貘、虎、豹、大熊猫、金丝猴、山魈等。苏门犀存在于大溪文化一至三期,梅氏犀存在于大溪文化一至四期,貘存在于大溪文化时期一至三期,大熊猫出土的最晚年代为遗址六期(中坝文化老关庙期),虎存在于大溪文化二、三期,豹存在于大溪文化一期,金丝猴存在于大溪文化一至四期,山魈出土的最晚年代为大溪文化四期。综上所述,大溪文化以后,大溪遗址并未出土有相关珍稀动物的骨骼。
动物物种的变化,除了这些动物自身适应能力之外,也与气候环境的变化、人类活动的过度开发等方面也有关联。
长江三峡西起于奉节瞿塘峡,自然地势上是我国二级阶梯向三级阶梯过渡地带,属于江河切割侵蚀区,从三峡峡谷至谷顶的高阶地,海拔高差数百米至千米,峡谷区域内地势陡峭,高山峰顶区域地势平缓开阔,因此在这个落幅变化很大的地貌单元内,形成了三峡极富个性的立体地貌景观。即在三峡峡谷内是中亚热带气候、植被,顺地势往上,梯级变化为北亚热带气候植被带和温带气候植被带,各个不同的气候植被带相应形成不同的自然景观,各个不同的自然动物群,分别生活在不同的自然景观内。三峡地区特殊的立体自然环境景观特点,为该区域的生物多样性提供了先决条件[23]。
苏门犀、梅氏犀、貘这类动物生存的气候环境为湿热气候下的森林、沼泽、河边、或热带雨林地区。虎、豹、大熊猫、金丝猴、山魈这类森林指示性极强的动物,对自然环境和气候环境的要求也很高。而该遗址新石器时代的“大熊猫——剑齿象动物群”恰巧说明这一时期的气候环境是适应以上这些动物生存的。新石器时代结束以后,气候环境发生变化,气候由原来的湿热气候逐渐转变为相对较冷的亚热带季风性气候,使得犀牛、貘等动物向更温暖更潮湿的南方迁徙。除此之外,由于人类活动的愈加频繁,尤其是农业的发展,先民改造自然的能力加强,地势较平坦的地方被开垦用作农田等,使得森林性动物的活动空间被破坏,生存空间被不断压缩,它们被迫逐渐远离遗址附近区域,迁往森林更为密集的地方生存。
2.家养动物
从出土的动物种类看,狗、猪、水牛是大溪遗址最主要的家养动物,各个时期均有发现,尤其集中在新石器时代大溪文化的一、二、三期。家养动物总量为1677件,占哺乳动物骨骼总量的15%(附表一、附表二)。
从出土时代上看,水牛出土的时代主要集中在一至四期,三期最多,其次为二期、一期、四期;家猪出土的时代主要为一至四期,三期数量占绝对优势,其次为二期、一期、四期;狗的时代主要为一至三期,二期最多、其次为三期、一期。从出土的数量上看,狗的数量最多,占家养动物总量的66%,家猪次之,占家养动物总量的19%,水牛最少,占家养动物总量的15%。说明遗址内已经进入到家养经济阶段,农业也应该有所发展。
大溪文化时期,狗的数量远远超过水牛和家猪,说明大溪先民对于狗的作用超于其他两种家养动物的作用。换言之,狗具备了肉食、农业需求之外的功能。遗址内出土有多个狗坑的遗迹,这应是作为随葬或祭祀的形式埋藏,这说明狗具有某种财富或宗教方面的特殊含义[24]。同时,狗还具备看家护院、辅助打猎等方面的作用,说明大溪先民对狗有着更为全面的物质、情感需求。
大溪遗址最核心的文化为大溪文化时期,出土动物骨骼最多的时期也主要集中在这一时期,尤其是一至三期,这与该遗址的文化特征极其吻合,说明该区域在大溪文化时期人类活动极为频繁。而历史时期出土的动物骨骼数量减少,尤其是家养动物的数量锐减,这说明该区域在历史时期的人类活动与新石器时期的活动方式有所转变。
五、结论
大溪遗址是三峡地区新石器时代晚期的重要遗存之一,具有重大的学术价值。出土的动物骨骼数量庞大,种群丰富,对这批动物骨骼的鉴定研究,可以让我们更为全面的了解三峡地区的古动物群、古气候等。
大溪遗址新石器时期生存有热带雨林气候的动物,说明那时气候比现在更为温暖湿润。出土物种的多样性是由其地理位置与气候环境决定的,三峡地区具有独特的立体地貌景观,长江作为重要水流资源,海拔高程越往上其植被特征越为显著,草丛、竹林、林缘灌木丛、阔叶林、针阔混交林、针叶林等。因此,遗址内出土的动物种类包括了内河淡水的水生性、喜水环境的草丛性、温暖潮湿的洞穴性、温暖湿润的森林性、温凉高冷的高山性等动物。
大溪遗址紧邻长江,淡水资源丰富,森林资源广布,具有丰富的肉食资源。新石器时期遗址出土的鱼类、爬行类等动物说明长江流域是大溪先民取之不尽的淡水资源;与此同时,数量庞大且种类丰富的野生动物是大溪先民用之不竭的森林资源。这些动物骨骼同时出现在大溪遗址,说明该遗址的渔猎经济发达,先民通过使用工具、分工合作、不断学习,掌握了高超的捕鱼技巧以及狩猎技巧。综上所述,大溪遗址的新石器时代,生业经济以渔猎经济为主,家畜饲养经济为辅。
处于长江较上游西侧的巫山大水田遗址,与大溪遗址相距28公里,该遗址也出土了大量大溪文化时期的动物骨骼。野生动物的种类与大溪遗址大致吻合,且也具备一定的家畜饲养经济,只是对于骨骼的利用程度明显高于大溪遗址[25],对于这一现象,需做进一步的研究思考。
处于长江较下游东侧的湖北秭归东门头遗址,时代包括新石器时代中期的城背溪文化、商代、周代、汉代、宋元、明清时期[26],其动物种群也具有明显的延续性。结合大溪文化时期的动物种群,在新石器时代的三峡地区,动物种类鹿科动物是绝对的优胜种,且种类多样;猴科动物也在该区域广泛分布,只是出土的种类略有不同;虎、熊、猪獾等这类森林性动物出土率较高;而普氏野马(东门头遗址)、爪哇野牛(东门头遗址)、圣水牛(东门头遗址)、貘(大溪遗址)、大熊猫(大溪遗址)、犀牛(大溪遗址)等这类已濒临灭绝的大型哺乳动物偶有出土,且数量极少,或许这些物种在新石器时代的三峡地区数量本身不多;水生动物(鱼类、软体动物等)的种群变化不大,说明长江流域的水生环境较较为稳定。商周以后随着农业社会的发展,三峡地区的野生动物种类除鹿科动物与鱼类外,动物种类以家养动物为主,说明这一时期三峡地区的社会处于同一发展水平,农业得以较大程度发展,家畜饲养占据重要地位。
附表一 大溪遗址各时期出土动物骨骼种类及其数量
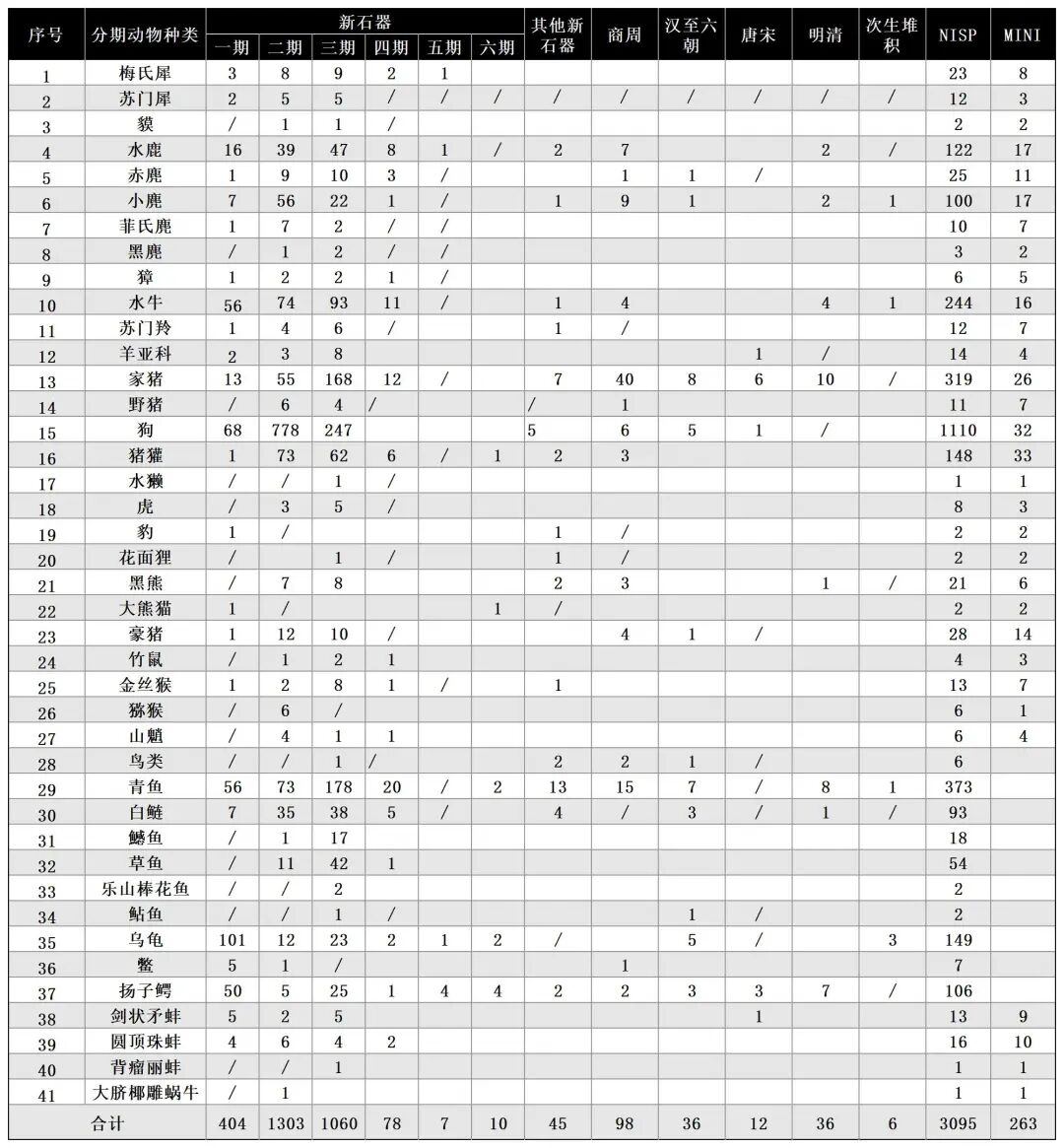
附表二 大溪遗址各时期动物最小个体数统计表

注释:
[1] 《重庆市巫山大溪遗址发掘报告》。
[2] 陈少坤、黄万波、裴健等:《三峡地区最晚更新世的梅氏犀兼述中国南方更新世的犀牛化石》,《人类学学报》2012年第31卷第4期,第381~394页。
[3] 同号文、武仙竹:《湖北神农架犀牛洞梅氏犀(真犀科,哺乳动物纲)化石》,《科学通报》2010年第55卷第11期,第1015~1025页。
[4] 陈全家、王善才、张典维:《清江流域古动物遗存研究》,北京:科学出版社,2004年,第156页。
[5] 西蒙·赫森著,侯彦峰、马萧林译:《哺乳动物骨骼和牙齿鉴定方法指南》,北京:科学出版社,2012年,第8~9页。
[6] 李凤、代玉彪、白九江:《巫山大水田遗址动物骨骼微痕分析与利用方式研究》,《第四纪研究》2004年第44卷第2期:第524~533页。
[7] 西蒙·赫森著,侯彦峰、马萧林译:《哺乳动物骨骼和牙齿鉴定方法指南》,北京科学出版社,2012年,第23~25页。
[8] 湖南省文物考古研究所:《永顺老司城遗址出土动物遗存》,科学出版社,2018年:第241~251页。
[9] 武仙竹:《长江三峡动物考古学研究》,重庆:重庆出版集团、重庆出版社,2007年,第139~148页。
[10] 李天元、武仙竹、李文森:《湖北郧县曲远河口发现的猴类化石》,《江汉考古》1995年第3期,第4~7页。
[11] 刘芳、常云平、张硕:《巫山蓝家寨遗址发现的金丝猴骨骼标本》,《重庆师范大学学报(自然科学版)》2014年第31卷第1期,第48~50页。
[12] 胡长康、齐陶:《陕西蓝田公王岭更新世哺乳动物群》,北京:科学出版社,1978年,第1~64页。
[13] 李天元、武仙竹、李文森;《湖北郧县曲远河口发现的猴类化石,》《江汉考古》1995年第3期,第4~7页。
[14] 李凤、白九江、代玉彪:《2014年度巫山大水田遗址动物骨骼鉴定初报》《西南文物考古》第一辑,第151~170页。
[15] 武仙竹:《长江三峡动物考古学研究》,重庆:重庆出版集团、重庆出版社,2007年,第49页。
[16] 黄万波、计宏祥:《江西万年仙人洞全新世洞穴堆积》,《古脊椎动物与古人类》1963年第7卷第3期,第263~272页。
[17] 林甲、李曼珍:《鲤鱼、鲫鱼、白鲢、草鱼的头骨比较解剖》,《哈尔滨师范学院学报(自然科学版)》 1964年1期,第127~156页。
[18] 莫林恒:《高庙遗址出土白鲢支鳍骨的鉴定与研究》,《湖南考古集刊》第9集,2011年,第260~278页。
[19] 周本雄:《山东兖州王因新石器时代遗址中的扬王舞遗骸》,《考古学报》1982年第2期,第251~260页。
[20] 侯彦峰、马萧林:《考古遗址出土贝类鉴定指南淡水双壳类》,科学出版社,2021年:第149~150页。
[21] 李凤、张七昌:《动物考古视角下的重庆古环境》,《重庆考古》公众号,2022-09-09。
[22] 夏武平等编著:《中国动物图谱兽类(第二版)》,北京:科学出版社,1964年,第79~90页。
[23] 李凤、白九江、代玉彪:《2014年度巫山大水田遗址动物骨骼鉴定初报》,《西南文物考古》第1辑,第151~170页。
[24] 邓晓、刘晓亮:《重庆巫山大水田遗址中独立“狗葬坑”初探》,《三峡大学学报(人文社会科学版)》,2019年第41卷第1期:第21~24页。
[25] 李凤、代玉彪、白九江:《巫山大水田遗址动物骨骼微痕分析与利用方式研究》,《第四纪研究》2004年第44卷第2期:第524~533页。
[26] 国务院三峡工程建设委员会、国家文物局:《秭归东门头》,科学出版社,2010年:第414~453页。
