秦岭 傅稻镰 | 不爱远航的稻农:中国沿海早期生业经济传统与海洋取向
字号:T|T
2025-02-25 09:16 来源:南方文物
摘要:长江下游是稻作农业早期发展和湿地稻田系统形成的关键地区。该地区新石器时代遗址的生计模式突出了淡水湿地对植物和动物食物资源的重要性。新石器时代晚期,从事稻作农业的农民主要依赖内陆地区(尤其是湿地和附近的林地)作为他们的主要蛋白质来源,而与海洋的联系非常有限。相较于旱稻和其他作物而言,水稻的产量更高,所以长江下游地区以水稻为主的生计策略可以支持本地人口的数量和密度快速增长。稻作农业需要大规模的劳力投入和对水资源的有效管理,而这会推动社会结构朝着更复杂的方向发展,位于长江下游地区的良渚文化便是一例。人口的增长可能在很大程度上被本地吸收,表明人口变化的主导趋势是向内集聚而非向外迁徙。朝鲜半岛、日本等地的其他农业扩散案例,进一步说明了稻作农业传播和人口语言扩散之间缺乏相关性。尽管稻作农业作为拉动因素推动了当地人口密度和社会复杂性的增加,却似乎并没有推动人群向外扩散。相反,从水稻到雨养稻作(旱稻)农业系统的转变或者稻作与其他雨养谷物的结合,更有可能推动了早期农民的向外迁徙和农作物的传播。
农业的出现对环境和人类产生了深远的影响。从Diamond到Ellis,各国学者对农业引发的各种变革进行着持续研究。宏观来看,农业在促进人口增长、扩大人口规模、形成语系和遗传谱系等方面均发挥了显著的作用。所谓“语言-农业扩散模型”强调农业的出现对人口的影响,认为其引发了人口增长和农业人口的外迁,并解释了大部分现代主要语系的地理分布扩散动因,如东南亚大陆和岛屿不同的语系的分布。因此,东南亚大陆的语言,如南亚语系,被认为是源于中国稻作农民向南的扩散;而主要分布在东南亚岛屿和太平洋地区的南岛语族似乎也是源于中国稻作农业的人口增长和向海洋的迁徙。Bellwood就认为,稻作农业起源于长江下游地区,那些将稻作农业经航海传播至台湾岛的开拓者,很可能是来自新石器时代的河姆渡等文化。
多学科的研究为这一设想做出了贡献。自20世纪30年代以来,考古学家就将台湾的物质文化与福建、广东和太平洋岛屿联系在一起进行研究。最早被认可的联系包括有肩石斧和绳纹陶器等器物。张光直的研究明确了台湾新石器时代文化的基本序列,包括与福建、广东地区考古学文化的关联。Bellwood和焦天龙等人一直倡导,稻作农业和海洋文化在约5000年前的新石器时代从长江下游地区沿海南下扩散,从杭州湾传到福建,最终到达台湾。与此同时,比较语言学的研究已建立起南岛语系与一些台湾先民语言最基本分支之间的关系。重建的原始语言词汇还确定了与农耕相关的词汇,包括稻和粟。最近,Sagart提出一个假设,即这些农业词汇的起源均可追溯到更早的汉藏语系或原始汉藏语言。正如这些语言学数据所暗示的那样,传入台湾的早期新石器文化传统中不仅包括稻谷种植,还包括粟(可能还有黍)等小米类谷物。事实上,最近在台湾南关里东遗址的植物考古研究证实了这三种谷物(稻、粟和黍)遗存在台湾地区的存在不晚于距今4300年,甚至可能早至距今5000年。
从20世纪70年代起,学界就将世界上地理分布最广的语系——南岛语系的语源追溯到台湾,这一语系所有基干分支都可以在台湾先民语言中找到源头。从中派生出了马来-波利尼西亚语系,而其他分支则传播到了东南亚岛屿、衍生到太平洋甚至马达加斯加一带。这棵语系树的结构催生出了关于人口扩张的所谓“快车道”模型,即从台湾出发,通过东南亚岛屿,最终通过拉皮塔文化扩张,在约3350年前传播至太平洋。尽管存在对这一语言模型的批评,但它仍然是解释南岛语系语言如何在历史上关联在一起的最为主流且最广泛接受的观点。
基于这个模型,台湾新石器时代的先民被认为是“原-南岛语族”。考古学家Peter Bellwood的一个重要贡献,便是综合了整个东南亚岛屿的相关考古证据,通过突出陶器等文化相似性,将印度—马来西亚新石器文化与菲律宾北部和台湾的文化联系起来。他根据各地语言模式和考古学文化进行分析,进一步发展了“语言-农业扩散模型”,其基本理念是:农业的发展和人们寻找新耕地的需求是人口扩张的主要动力。随着这些不断增长的农业人口向岛屿扩散,他们在很大程度上取代了,或一定程度上融合了原有的狩猎采集人口。目前,有关人群迁徙至台湾和稻作农业在台湾以外地区传播的相关植物考古学证据仍然有限。在岛屿环境,农业似乎发生了一个重大变化——芋头和山药等根茎类作物变得比水稻更为重要。尽管缺乏有关农业的直接证据,但在全新世晚期的东南亚岛屿和台湾地区范围内,这个由南岛语族农人和水手组成的不断扩张的“新石器化格局”统一了语言学和考古学的叙事。
对于这样的历史叙事,我们可以提出三个问题。首先,为什么是稻属植物?为什么稻作农业成为影响人口增长和农人迁徙的决定性因素?那么其他种类或形式的粮食生产是否也能成为人口迁徙和增长的推动力?其次,我们想知道:是具体哪种稻属品种,旱稻还是水稻?稻作栽培方式较多、分布范围很广,有从山地的刀耕火种到密集的灌溉系统等。那么在这些不同的生产策略中,哪种栽培方式下的稻属作物推动了人群向台湾及其他地区迁徙?尽管这个问题受到的关注相对较少,但大多数研究倾向于认为是更加密集型和高产的水稻栽培。事实上,我们的研究表明,目前的证据和逻辑推断均与此主流观点相左。最后一个问题则是,基于目前所谓稻作农业及其扩散的考古学证据,我们是否能够得出稻农远航形成南岛语系的结论?
针对以上三个问题,我们认为:早期从事稻作农业的农人并没有表现出特别的扩张性,也没有进行过多的海上远洋活动;相反,他们更倾向于专注对淡水湿地的开发和利用,很少与海洋有所接触。从长江下游地区的植物考古、动物考古和聚落考古的证据中,可以清楚地看出这种倾向。事实上,高产的水田稻作农业推动的是人口在单位面积内的增长聚集而不是地理范围上的扩张。除了长江下游地区的稻作证据外,还需要考虑和比较其他形式的粮食生产,包括旱作小米类谷物、低强度的旱稻品种和蔬果栽培。实际上,当考虑到潜在产量、劳力需求、土地投入和可持续性时,旱作农业(包括小米类和较低强度的旱稻)更可能推动地理范围的扩张以便农人寻找新的土地来满足人口的增长。结合沿海的狩猎-捕鱼传统,目前看来新石器时代长江下游地区从事水田经济的稻农与传播到台湾、东南亚地区的农业形态及相应的人群迁徙之间没有任何联系。因此,已有的假设需要被否定或加以修正。
长江下游一带是稻作农业早期发展的关键地区,广义上包括环太湖地区和宁绍平原(图一)。一直以来,在对新石器时代稻作农业和南岛语族的起源传播研究中,该地区的河姆渡、马家浜等新石器文化都占据重要地位。然而,近年来中国各地涌现出越来越多的新石器时代相关发掘成果,学者们发现了许多很早便有稻作栽培实践的考古学文化,诸如长江中游的彭头山文化、汉水流域的八里岗遗址前仰韶文化、淮河流域的贾湖文化和顺山集文化等,它们很可能说明了存在多个独立起源发展的稻属作物驯化过程。不过,由于长江下游地区在地理位置上离福建沿海和台湾最近,又有相对最完整的考古学及植物考古证据链,因此仍然是很多学者探究淡水资源和海洋资源在稻作农业演进中不同作用的重点地区。
1.跨湖桥 2.河姆渡 3.田螺山 4.马家浜 5.草鞋山 6.绰墩 7.澄湖 8.姜里 9.良渚古城遗址 10.茅山
在长江下游地区,通过对农业和非农业生计、技术、景观和饮食的研究,已经可以梳理出稻作农业兴起相关的文化演进过程。特别是在过去二十年里,该地区的考古成果日益显著。水稻驯化已经可以通过多种性状特征的变化来进行探究。例如,与野生祖本原有脱粒性相对的非脱粒性是其演化出的依赖人类收割的驯化特征,从脱粒到非脱粒的迅速转化出现在约7000至6000年前,正好对应着河姆渡和马家浜文化的存续时间。而水稻扇形植硅体,实际上在大约6000年前也已经出现了尺寸上的线性变化,这表明水稻(从叶片的角度)在被驯化过程中持续演化。这种变化与谷粒尺寸的演变相一致,种子随着驯化完成及之后的持续发展变得更为饱满。此外,在距今6000年之后,稻谷继续分化为短粒和长粒两种类型,并分别成为不同聚落和地区相对稳定的品种。这些不同的水稻驯化谱系很可能最终稳定为今天的热带和温带粳稻。尽管现代温带粳稻的一些特征可能是更晚阶段演化而来,但这种粒型上的差异在新石器晚期长江下游的不同区域和聚落中就已经显现。
驯化稻各种性状特征演化的速度和时间点与农业技术的发展密切相关。最初的驯化可能是通过土地管理和特定种植与收割技术的长期作用来实现的,在这一过程中,人类行为与植物演进具有相互依赖性,产量与人为的播种收割共同进化。Allaby等人最近的研究估计,狩猎采集人群对稻属植物最终导致驯化的利用,始于距今约13000年。而在8000至6000年前,水稻演化的速度明显加快,这一阶段也通常被认为是水稻的驯化期。而最早的水田遗址可以追溯到这一阶段的末期,马家浜文化晚期(6000—5800年前)的多个遗址均有相关发现,如草鞋山、绰墩(图二)和姜里遗址等。
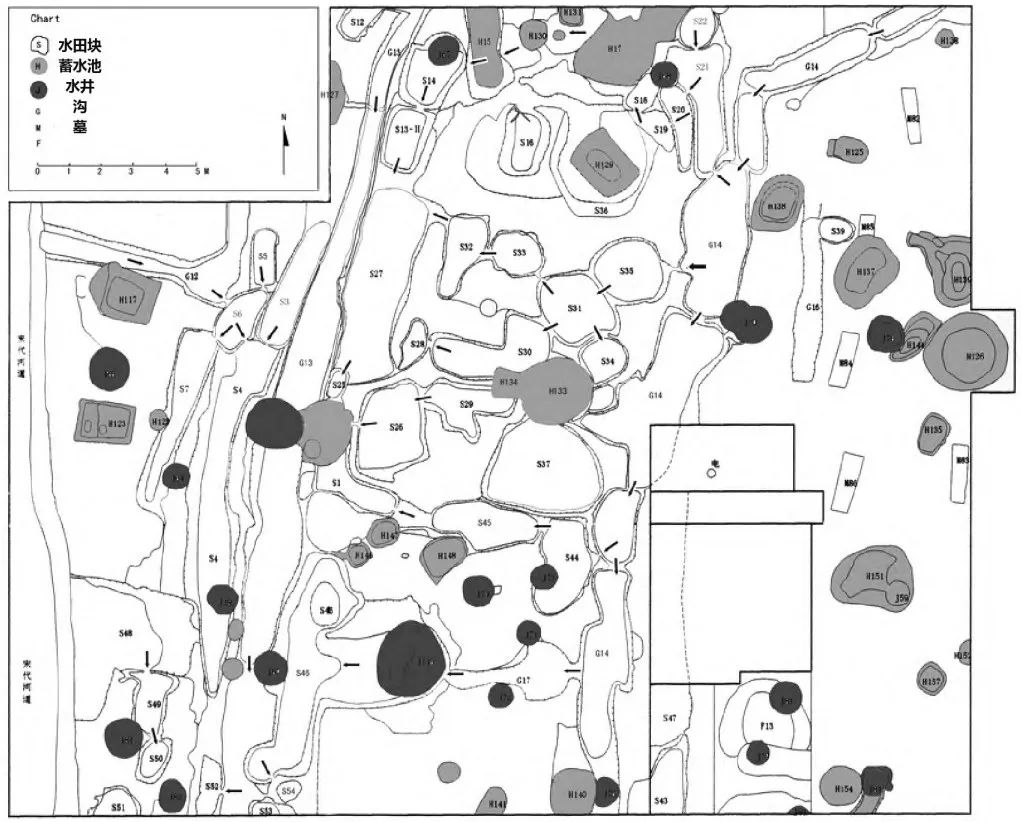
图二 绰墩遗址水田遗迹(Fuller et al. 2009)
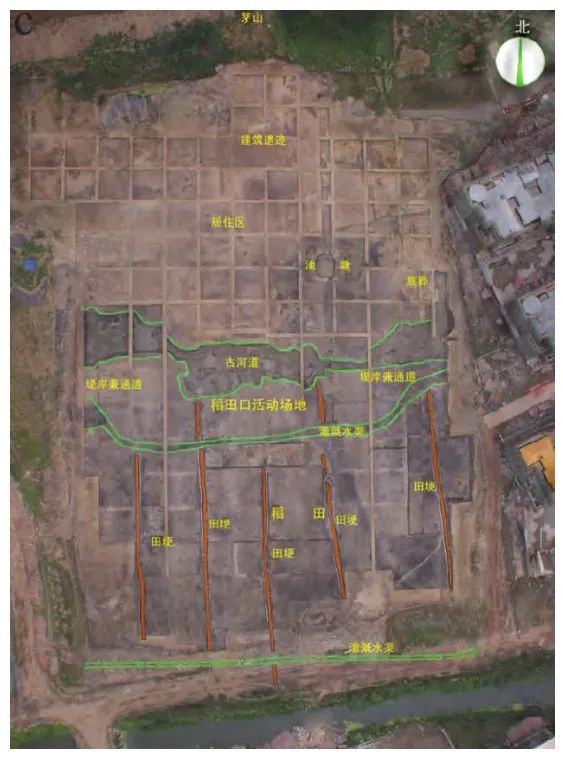
在受控的农田环境中,可以预期对水稻的形态特征产生更强的人工选择(例如生长习性和叶片形状),而在农田中相对独立的持续生长演进也有助于形成不同区域间呈现谷粒形态分组分布的特征。最早的水田是非常小的浅坑状遗迹,直径通常在1~2米之间,总面积不超过10平方米。这种耕作策略的一个好处便是能通过严格控制水源和排水来改变水稻野生祖本“多年生”的原有特征,从而实现更高的年产量。随着时间的推移,水田的面积逐渐扩大,从崧泽文化一直延续到良渚文化早期(约5500—4800年前)(图三)。然后,在良渚文化晚期阶段,出现了全新的水田系统,它拥有系统化的灌溉、排水和精心规划、规模庞大的布局(图四)。在浙江临平茅山遗址发现的位于大型水田遗迹之下的早期浅坑状水田遗迹,清楚地显示出良渚文化中晚期阶段社会生产向更加集约化的稻作农业转变的趋势。
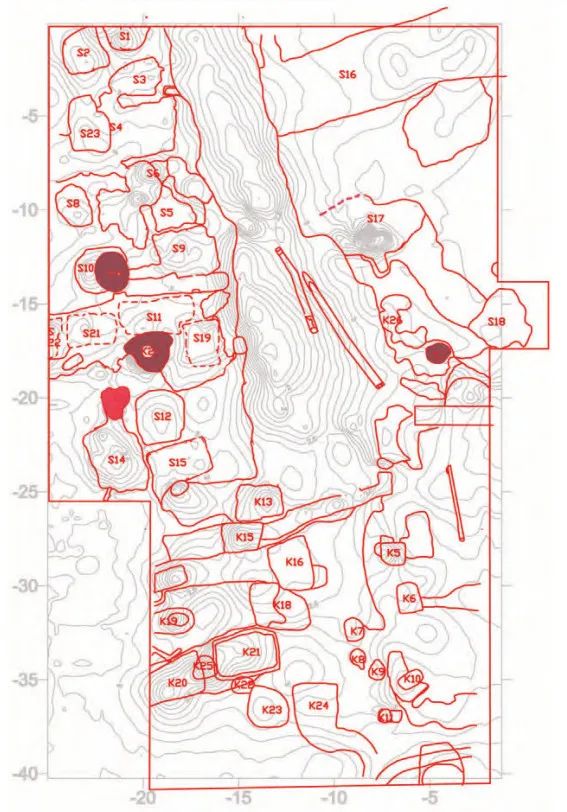
(良渚晚期,Zhuang, Y. et al. 2014)
除水田系统的明显演化外,有关农具的考古学证据也揭示了清晰的演变轨迹。在水稻完成驯化的最初阶段,我们并没有发现大量明确跟收割相关的工具,用来收割和耕作农田的“工具套”主要出现在产量大幅提升的驯化后阶段。长江下游地区新石器时代晚期的农具包至少包括三角形“石犁”和带柄“破土器”,可能均用于翻动农田土壤;称为“耘田器”的石刀,用于手工收割单个稻穗;还有用于割断稻秆的更大型的石镰刀。三角形石犁头和其他收割工具类似,最早出现在约5500年前的崧泽文化晚期。这些工具在长江下游的新石器时代和青铜时代逐渐发展,特别是石犁类工具尺寸日益增大,最终被铁器所取代。它们的重要性在于其反映了水田经济生产与所需大量劳动力投入的关系,使人口与高产且重要的稻田紧密联系在一起。
水稻是长江下游地区在新石器时代唯一驯化栽培的农作物,目前尚未有考古证据表明粟黍在该地区被栽培和食用。不过,其他淡水湿地植物始终被先民们开发利用,其中占重要地位的有芡实和菱角,而林地中的橡子等坚果在水稻被驯化之后则逐渐退出历史舞台。最新对田螺山遗址出土菱角的形态学分析显示,菱角也可能已在距今7000年被栽培驯化。尽管坚果和果实类林地资源也被史前先民利用,但水稻、菱角和芡实在这一地区的可食用植物遗存中占据绝对的主导地位,凸显了淡水湿地资源对生业模式的重要性。
湿地的关键作用也在田螺山和跨湖桥遗址的动物遗存中体现出来。经鉴定,鸟类骨骼主要来自湿地品种,如鸭科(Anatidae)、雁亚科(Anserinae)、秧鸡科(Rallidae)、鹭科(Ardeidae)和鹤科(Gruidae)。虽然鱼骨遗存的发现和研究相对较少,但田螺山有一项样本量很大的分析可供参考。该研究对湿筛样本中的174340块鱼骨进行了鉴定和分析,揭示出淡水湿地鱼类在其中占主导地位,如鳢鱼(Channa)、鲤鱼(Cyprinus)、鲫鱼(Carassius)和鲶鱼(Silurus)。这些物种都可以生活在稻田周围或附近更深的水域,也正是可供菱角或芡实生长的地方。根据鱼骨尺寸的分析复原,人们会全年捕捞鲤鱼和鲫鱼,而针对鳢鱼则更多在春季进行捕捞。鱼类组合还显示,人们对少量(0.7%)海鲈鱼存在一些沿海或河口的捕捞活动,尽管这种鱼在不繁殖时也会游入淡水河流。虽然考古学者在田螺山遗址采集到若干大型金枪鱼椎骨,在跨湖桥发现了一块海豚骨骼,但海洋和沿海食物资源仍属个例,不能被解释为新石器时代该地区先民生活的日常。因此,这些稻农主要依靠内陆,尤其是湿地的资源,作为他们主要的蛋白质来源。
大型哺乳动物包括各种鹿类、一些猪和水牛,同样反映了一个利用湿地和内陆林地资源的环境。大量獐(Hydropotes inermis)和圣水牛(Bubalus sp.)的骨骼遗存显示先民在湿地及其周围进行主要狩猎活动,而梅花鹿和马鹿(Cervus spp.)则指向林地环境。少量的家猪和野猪骨骼(Sus scrofa)被认为是狩猎野猪和对猪的早期管理在8000年前后同时存在的证据。而来自良渚文化出土器物上的动物形象,同样强调了湿地动物和鸟类等资源(图五)。
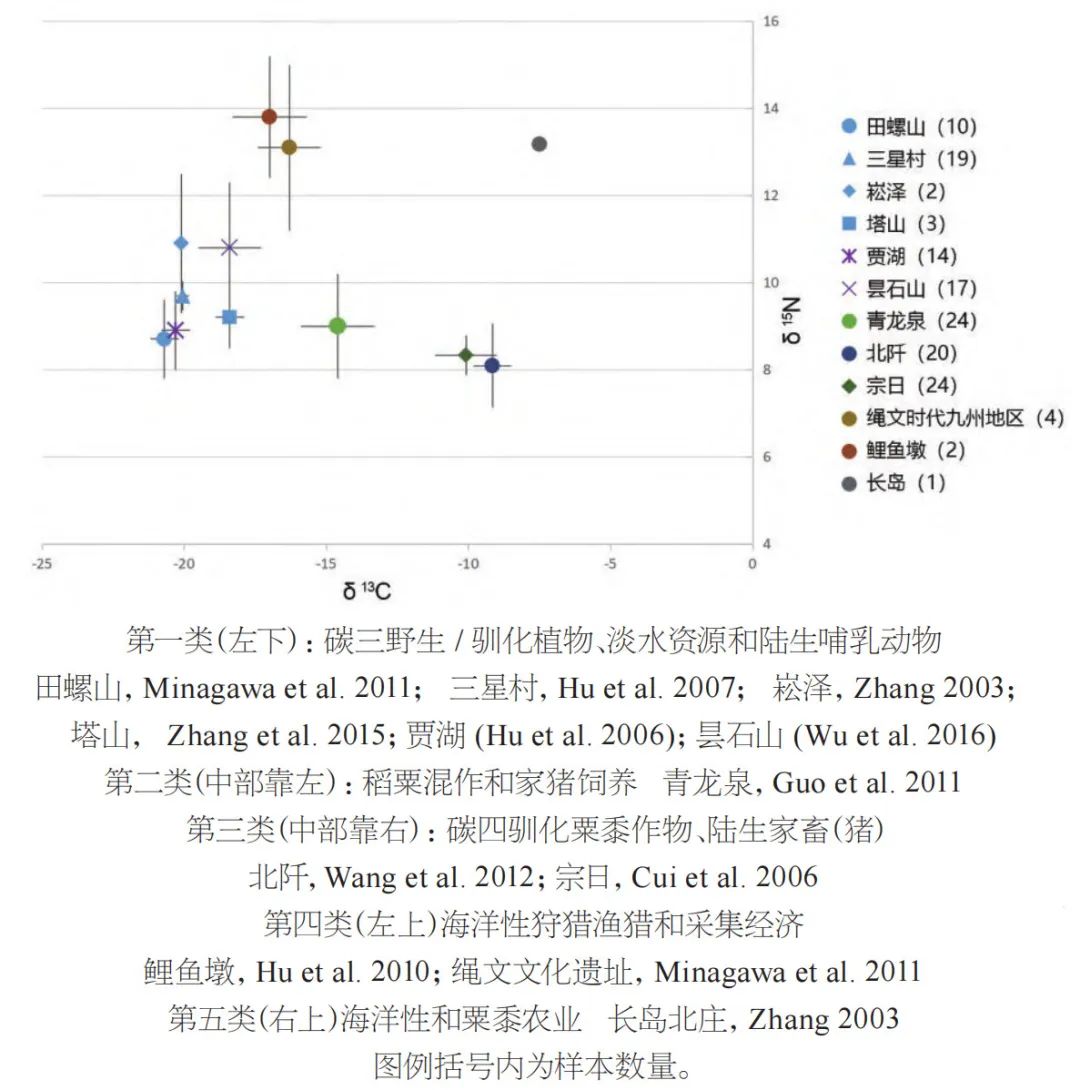
综合来看,依据长江下游地区新石器时代遗址发现的食物资源证据,我们可以重建该地区的早期土地和资源利用情况。物质文化中的图像也反映了相同的资源利用模式,其中鸟、淡水的鱼和龟鳖是反复出现的主题(图五)。当地人群的环境资源利用模式也可通过稳定同位素分析数据体现在食性中。从数据分析,长江下游地区先民以C3陆生和湿地类型食物为特征,其稳定同位素数据的分布明显不同于依靠海洋资源为生的狩猎采集类型、海洋资源加粟黍C4类农业的类型和单纯的内陆旱作C4小米类农业类型(参见图六)。值得一提的是该区域目前还发现有两处独木舟遗存,分别来自跨湖桥遗址(8000年前)、茅山遗址(4500年前)(见图七),显示了简单的河流湿地舟船技术的存在。
图六 碳氮稳定同位素体现不同食性和经济模式
(浙江省文物考古研究所等:《跨湖桥》,文物出版社,2004年)
因此,我们得出结论,新石器时代长江下游地区的先民对海洋既无谋生目的,也没有交通技术上的联系。相反,他们主要依靠淡水湿地和附近的林地资源为生。这些先民似乎将目光更集中地投向了内陆,而非海洋。
三、水稻与其他作物生产体系:人口统计和土地利用模式的比较
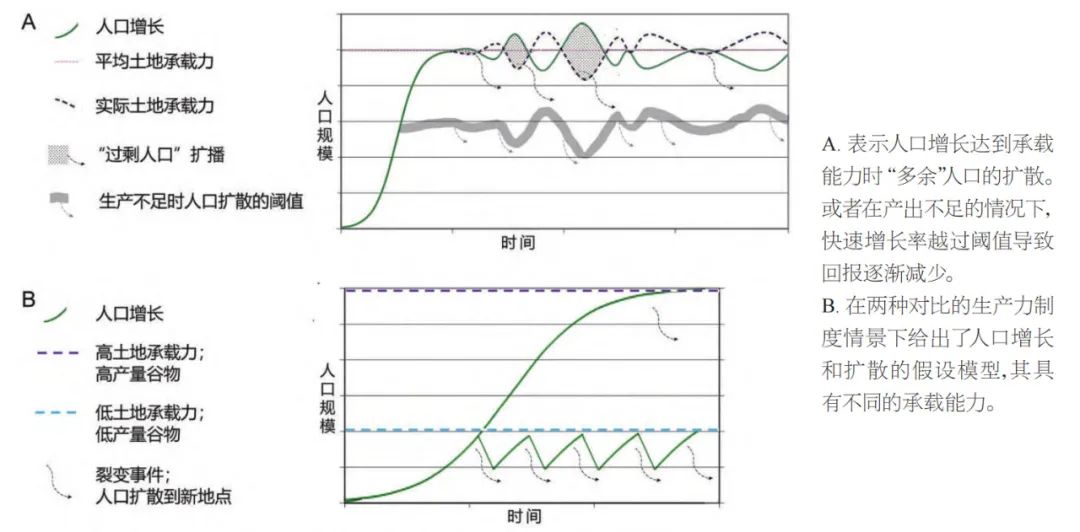
稻农从长江下游往外迁徙的观点是基于人口扩散的底层逻辑。这个理论假定,人口不断增长导致人群分裂,子群体向外迁移寻找新的土地进行定居和农耕。Rindos解释说,当地区人口增长到达或超过其自然承载能力时,此类迁移事件便会发生。农业社会的土地承载能力本身会因为收成的变化等因素在不同年份间波动,而这种不稳定性的程度可能会加速或减缓人群的总体迁徙速度。Shennan团队最近对欧洲新石器时代的考古年代数据进行了综合分析,通过人口统计学的分析方法指出,扩散趋势往往发生在地区人口快速增长但增长减缓之前;换句话说,早在达到承载能力之前人群便开始寻找新的农业领地。因此,欧洲的数据表明,人口不仅在达到极致规模时可能通过迁徙寻找新的农业领地(如Rindos模型所示),而且在一个快速增长的中间阶段这种迁徙扩散也会发生。
从比较民族学研究的角度来看,这个理论也是合理的。许多传统的小规模社会在低于其承载能力的情况下运作良好,正如萨林斯Sahlins所称的“生产不足(underproduction)”或“资源低效利用(underuse of resources)”。通过各种传统生产体系的数据,包括人口和计算得出的潜在生产能力,萨林斯发现,这些社会全都是低产即生产不足的。只有几个群体的真实生产力达到了预估值65%或75%的水平,而平均生产率仅为预估值的45%左右。因此,可能并不是土地承载能力本身推动了人群的分裂,而是人口增长达到一个阈值时,需要投入更多的努力来供养更多的人口。但无论如何,总的潜在承载能力的确会影响人口增长的速度以及迁移开始的时机(图八)。
这些观察引发了我们关于东亚和东南亚先民早期生计方式的两个问题。首先,不同地区或作物的承载能力(CC)及其相关的资源利用不足(~60%CC)之间是否存在特定和固有的差异,这些差异会否提高或降低人口增长的上限?其次,是什么样的差异决定了子群体开始扩散的时机?现有证据表明,与其他雨养农业形式(包括雨养稻、旱稻及小米类)相比,水稻栽培似乎不太可能推动人口的迁徙。
众所周知,水稻产量在其生长季内受水资源的影响而变化显著,同时,对劳动力投入的需求也存在差异。我们曾提出一个观点,即水稻农业对劳动力组织投入的需求较高,这可能会限制一些社群在有其他资源的前提下投入水稻栽培的吸引力,甚至可能存在一个社会复杂性的阈值,低于该阈值的社会群体会避免以水稻农业为生。更重要的是,就潜在的承载能力而言,也可以根据养活一个自给自足村庄或典型新石器聚落所需的土地来估算这种固有的差异。为了估算新石器时代农业聚落所需的可耕地面积,我们汇集了一系列关于水稻、旱稻和传统小米农业每公顷产量的民族志和古文献数据。将这些数据转化为卡路里产量,并考虑到人均每年消耗的谷物作物数量(假设是以谷物为主食)以及聚落人口数量,可以初步估算出社群所需的耕地面积。当然,我们提供的是一个大致的数量级,这些对人口估计并不是绝对精确的。
对于人口规模,我们以考古遗址的大小为依据并结合先前的人口估算成果。如长江中游的湖南澧县城头山遗址(距今6500—6000年)面积约为8公顷,长江下游浙江余姚河姆渡遗址(距今7000—6300年)面积约为4公顷,同时期的余姚田螺山遗址(距今7000—6300年)面积约为3公顷等。所有这些遗址都有可信的最大面积调查数据,即它们发展鼎盛时期的规模。中国新石器时代这一阶段聚落的人口密度大约认为是每公顷50人,比如基于河姆渡遗址的建筑数量和面积就有过这样的估算。基于仰韶文化早期姜寨遗址房屋面积和墓葬数量的研究显示,北方这一阶段小米值农业聚落的人口密度为每公顷53.5人。
很多研究提供了近现代每人每年消耗水稻的大约数值。根据Grist的说法,每人每年需要约250千克未去壳的稻谷来满足每天约2000卡路里的热量需求。在传统东南亚地区,估计值则为每人每年160千克。印度奥里萨邦沿海地区传统饮食的典型摄入量也是大约每人每年160千克,假设该数据代表去壳的稻米,其重量则相当于未去壳稻谷的60%~70%。这些对当代水稻摄入量的估算可能占人们总热量摄入量的80%左右,而河姆渡、跨湖桥、城头山等遗址植物考古学证据显示,新石器时代先民的饮食更多样化。当时的饮食富含其他碳水化合物,比如橡子、菱角,在城头山可能还会有极少的一些粟。因此,我们假设在新石器这一阶段先民的饮食中,水稻占总摄入量的50%左右(如果像现代饮食习惯一样,谷物占总饮食的75%~80%,那么土地需求的估算将提高50%~60%)。
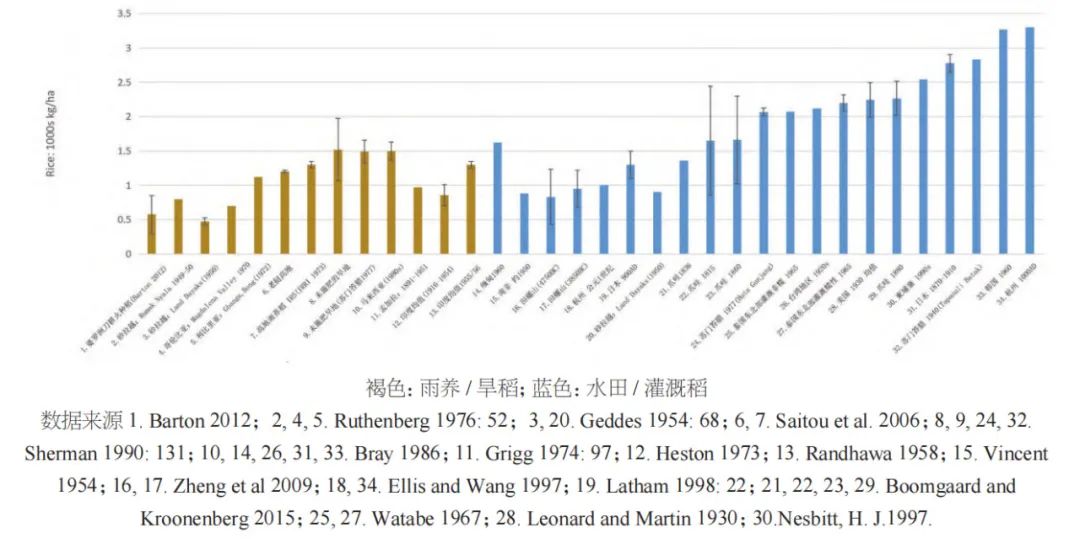
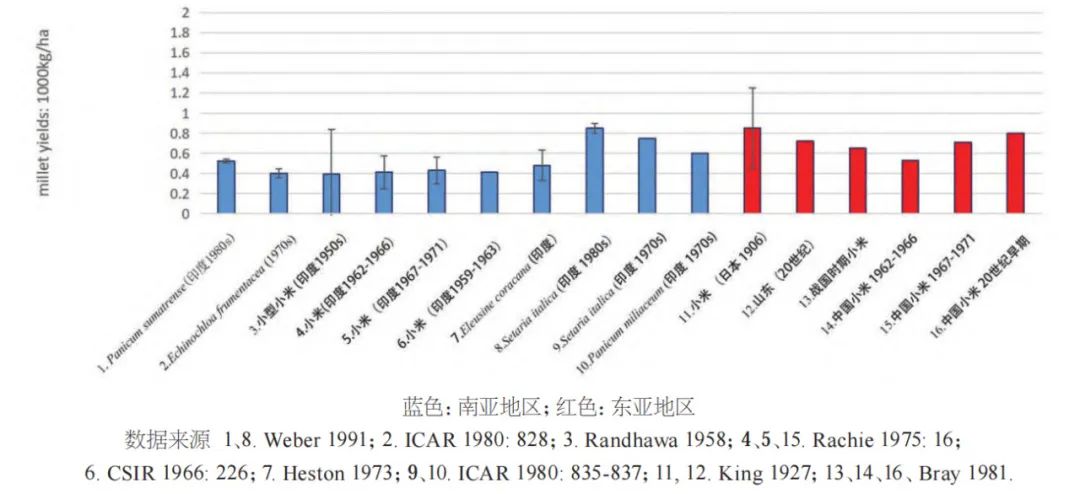
古代的作物产量很难估算,因为它直接取决于土地利用系统。而现代传统农业的产量不能直接借鉴作为史前时期的模拟数值。一般而言,水田稻作的产量肯定要比雨养稻高,因此,合理产量的下限可以基于旱稻生产力的数据。旱稻的产量范围从每公顷约480千克到当代某些产区的1500千克不等(见图九)。我们收集的旱稻比较数据平均值为1062千克每公顷,但若仅考虑巴拉望岛和婆罗洲的数据,平均产量仅为578千克每公顷,最低产量甚至只有229千克。水稻的平均产量汇总数据的平均值为1897千克每公顷,传统湿地水田的产量下限约为1500千克每公顷。而史料记载显示10世纪的日本水稻产量约为1300千克每公顷,约2000年前的汉代杭州地区观测到的水稻产量约为1000千克每公顷。据此,按新石器时代田螺山遗址(距今约6700年)水田遗迹中水稻叶片的植硅体密度估算,该地区早期未改良的湿地稻谷产量估计为830和950千克。据上,我们保守起见,将估算值调整为800或900千克,结合现代人群消费差值,便可大致确定中国新石器时代一些代表性遗址所需的水稻种植面积(见表一)。
表一 长江流域水稻消费、土地需求和承载力估算
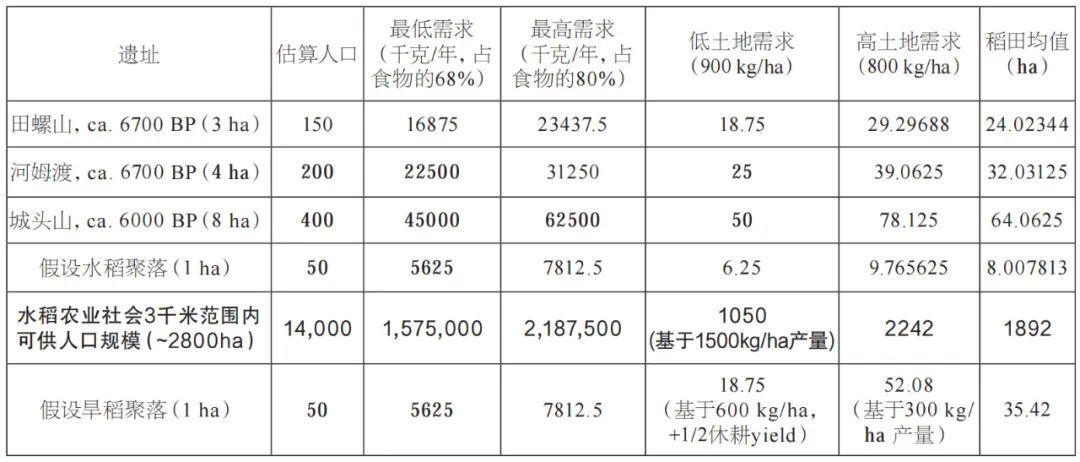
根据以上计算,我们估计新石器时代水田农业聚落,每公顷居址面积(或每50人)需要6.25至9.75公顷的水田耕地,中位值大约为8公顷(见表一)。我们的产量估计也相对较低,这意味着如果每公顷产出1000千克或更多的稻米,则每人所需的土地将更少,而当地的承载能力将超过我们现有估算。民族志及史料表明,大多数农田位于定居村落附近的3千米范围内,距离超过4千米的农田几乎不可能存在,因为先民需每天步行往返于农田和村舍家中。这表明大约2800公顷的土地可以轻易支持约14000人的人口规模。
水稻的高产量与雨养稻和小米类农业形成了显明对比(图九~图一〇)。东南亚地区传统的雨养稻有很好的文献记录,Barton总结指出婆罗洲的雨养稻作产量相当低,约为229至1000千克/公顷。对于新石器时代的旱稻生产来说,其产量也相应的大约可估算为水稻的一半,即每公顷约为400至500千克。这一低产量还会受到田地肥力下降和杂草干扰的影响进一步恶化,从而不得不采取轮种的方式。除非可以利用外部肥料补充地力(如家养牲畜的粪肥),否则现代传统雨养稻通常也还是采取轮作制种植。例如,在菲律宾伊班人的传统农业中,每年每人需要开垦约0.33公顷的稻田,而一个由140人组成的长屋村庄每年需要50公顷的耕地。根据这些数据,伊班人在一个定居点最多可居住14年便需要搬迁,而考虑到区域内某些土地的不适宜性以及社群年龄性别构成的变化,10年被认为是更好的估算。
因此,伊班人的土地需求大约是长江流域新石器时代聚落估算值的4倍(见表一)。这意味着以雨养稻为基础的定居点的土地承载能力大约是湿地水稻经济的四分之一。假设人口呈匀速增长,那么基于农业人口迁徙的底层逻辑,旱稻农民的人群分裂迁徙以寻求新空间的频率大约就是水稻农民的4倍。考虑到旱稻农民还需要为休耕而转移田地,或者干脆需要全部搬迁(例如像伊班人这样的村落每10到15年搬迁一次),具有流动性的文化传统和不断建立的新定居点可能会鼓励形成一种迁徙的固定模式。而与之相反,随着时间的推移,湿地水稻生产力的增加使得水田经济支撑的人口得以更加密集地聚居。
相比之下,中国北方传统的粟黍农业虽然每年产量较低,但黄土的潜在肥力也可以消减对田地轮作的需求。图一〇说明了各类小米可能产量的范围,由于对比数据较少,我们只包括了粟、黍和印度的各类小米。我们还假设早期小米产量的差异不是很大。例如,基于相同的实验条件,印度的实验发现黍的平均产量仅略低于粟(可能达到粟的95%)。正如何炳棣研究显示,中国北方的黄土具有含量很高的矿物养分,主要受到的是不同作物水分吸收潜力的限制。何从对周代农业记载的文献出发进行演绎推断,周人可能在第一年清理开垦土地,第二和第三年种庄稼,然后休耕一年。基于这种轮作方式,我们估计在黄土最肥沃的地区,每50人需耕种30~36公顷的土地,约为长江下游湿地稻作所需的4倍。一个具有这种生产力水平的3千米范围内可能支持4000人规模的聚落,但需要考虑典型的新石器时代小米农业土地承载能力会更低,可能只接近其一半。例如,水源贫瘠又缺乏规模灌溉的土地可能需要每两年休耕一次,增加土地需求并降低了承载能力。而随着小米种植从黄土高原扩展到亚热带和热带肥力较低的土地上,可能每三年中有两年或更多时间用来休耕。因此,随着小米栽培扩散到黄土高原以外的新社群,为了保持相同的生产力水平,就需要越来越多的可耕地面积。
基于耕作系统的性质,我们可以得出结论:长江中下游地区以水稻农业作为主导生计策略可以支持当地人口保持高密度和不断增长的态势。通过扩大和强化生产,人口增长主要在本地被吸纳。从这个意义上说,湿地水稻农业的发展推动了更大规模、更集中的人口聚居地的形成,这些地区的单季农业模式还可促进社会分工和专门化,支撑那些从事玉石器加工、陶器生产或宗教仪式活动的非农业人群。这个过程的最终结果是在长江下游的良渚文化和长江中游的屈家岭-石家河文化率先出现超大规模的城市型聚落。这两个区域文化都依赖本地大规模的水田经济作为基础,临平茅山遗址发现的水田遗迹就是实证之一。综上,新石器时代以水稻为主要作物的农业聚落中,人口会朝着更加密集的趋势增加,而非向远方迁徙。
水田稻作带来的较高人口密度既是湿地经济的产物,也是湿地环境开发的推动者。因此,长江流域比如太湖地区的湿地景观包括由稻田开发而扩大的自然水路交通网络,形成一个更利于鱼类资源利用、运输和社交的环境,也利于更大规模可持续的人口增长。成规模可持续的水田经济需要更多劳动力投入,由此产生的社会和经济组织形态也对进一步的社会和政治发展起到关键作用。这样的经济基础提供了一个很好的背景,帮助我们理解这一时期为何会出现良渚古城遗址这样的大型水利工程体系。这样的公共工程系统有助于良渚社会经济的发展,包括特有的贵族手工业经济玉器生产体系,适应于更高水平水稻农业的专门化农具生产体系等等,促进了区域社会复杂化的进程。
图一〇 传统及历史时期不同小米类农业的产量比较
接下来讨论三个东亚地区农业往不同方向扩散的例子,与上文所述的长江中下游水田经济与复杂社会发展形成对比,可以进一步说明湿地稻作农业与稻农扩散传播之间缺乏关联性。
1.作为补充的水稻:东北亚地区的早期农业与海洋文化
朝鲜半岛、日本列岛等地所在的东北亚地区,农业起步较晚,并从中国引进了主要的农作物。粟黍类小米和粳稻作为农作物从中国传播到朝鲜半岛,之后又传至日本。朝鲜半岛出土的小米类遗存可追溯到距今5500到5000年的栉文时代中期。同时期的小米遗存在俄罗斯远东地区滨海边疆的遗址上也有发现。稻米抵达朝鲜半岛的时间更晚,可能在距今3500年左右,相关年代数据仍有争议。
东北亚地区谷物栽培和农业技术的引入很可能伴随着农民迁徙,根据考古学的证据,这一传统起源于中国东北部。同时,近年来一些历史语言学家主张将朝鲜语和日本语追溯到一个假设起源于中国东北的跨欧亚语系“Transeurasian”。值得注意的是,这波农业化是由小米类较低生产力水平的谷物所驱动的,而不是湿地稻作。水稻传播一般认为是通过山东半岛传播至辽东半岛,再通过朝鲜半岛沿海聚落南下最终传播到日本,它是已有小米类农业经济在东北亚地区出现千余年后的“附加(add-on)”作物品种。并且山东半岛和辽东半岛的植物考古证据显示,稻作东传的中国青铜时代,这些地区自身的水稻栽培也并未占据主导地位。在韩国青铜时代的农业中,水稻作为选择性采用的作物品种,与小米、大豆和其他作物并存。此外,早期栉文陶器时代的贝丘遗址体现出对海洋食物资源的重视,显示出在农业文化传入之前,该地区海洋技术利用和发展的普遍性。实际上,在朝鲜半岛栉文时代晚期和无纹时代,海产品仍是人们生计活动的重要组成部分。
因此,农耕在朝鲜半岛被接纳和发展是循序渐进式的。从采集为主到出现农耕文化的转变确实可能代表了一种农业扩散现象,这也与朝鲜语和日本语的祖先以及跨欧亚语系的假说相关联。然而,无论是水稻还是旱稻,都是后来被当地小米农业、采集和海洋渔猎人群所采纳的一种附加作物,而不是文化或人口变化的经济驱动力。
台湾地区的农业起源问题,与福建沿海地区的情况相互关联。长期以来,学界普遍认识到台湾岛、附近的澎湖群岛和福建沿海的史前文化之间存在着密切的联系和互动。从晚更新世到约6000年前,台湾岛的先民处于无陶器的“旧石器时代”,第一个制陶文化被认为是大坌坑新石器文化。许多学者认为,大坌坑文化反映了原始南岛语族人群(Proto-Austronesian speakers)从粤东及珠江三角洲等地迁徙抵达台湾岛的时间。例如,早至6800年前,珠江三角洲地区就出现了树皮布石拍和拔牙习俗,可能与台湾后来的相关文化传统存在联系。在珠江流域的一些遗址中,发现了加工各种块茎、棕榈和其他野生淀粉类植物食物的证据,这表明在稻作引入之前(约4600到4400年前),该地区以采集为生,也可能有园艺类种植(vegeculture)。而靠近台湾的福建沿海地区,大量的贝丘遗址印证了先民对海洋鱼类和沿海贝类资源的利用,狩猎动物资源中未见家猪的相关证据。作为台湾第一个陶器文化,大坌坑文化延续了类似的海洋和沿海资源利用传统,珊瑚的利用在澎湖群岛和台湾的遗址中也都可以看到。这些发现表明这些沿海地区和岛屿的先民非常重视对海洋资源的利用。
在距今约5000到4500年的大坌坑文化最晚期,台湾西南部出现了最早的谷物利用证据,包括来自南关里遗址的稻米及南关里东遗址所出的稻米和各类小米。最近发表的植物考古成果证实了南关里东遗址出土大量的粟(Setaria italica)、黍(Panicum miliaceum)、稻米和金狗尾草(Setaria pumila,又称S. glauca auct. pl.)。小米在这个组合中占主导地位,并且根据出土区域缺乏粘性土壤或田地系统的迹象,推测这些同小米共出的稻属植物可能是雨养稻。距今4500年之后,台湾出现了四个地方性的新石器时代中期文化。台湾南部富山文化潮来桥遗址最新的植硅体证据证实了这里至少约4200年前便存在驯化稻。因此,该地区可能为前往菲律宾群岛航程的始发地,稻谷和小米栽培或许由此引入菲律宾北部的吕宋岛。
在福建省北部,最近的植物考古数据显示距今约4500年已存在稻粟混合农业。位于内陆山地丘陵地带明溪县的南山遗址包括有一系列洞穴居址,其年代可追溯到5000至4400年前(表二)。尚未完整发表的植物考古数据表明,存在稻米和粟黍两种小米。此外,最近在闽侯县白头山遗址的发掘(表二),木炭测年结果为距今4800—3700年,也发现了水稻和黍的植硅体证据。位于福建省沿海地区的黄瓜山遗址(距今4500—3900年)和屏风山遗址(距今3800—3400年)都有栽培稻的直接碳十四测年结果(表二)。尽管稻米占植物遗存的主导地位,但这些遗址的炭化大植物遗存和植硅体证据都清晰显示了稻米和粟黍的混合作物模式。
表二 本文涉及的华南、东南亚史前遗址出土谷物情况
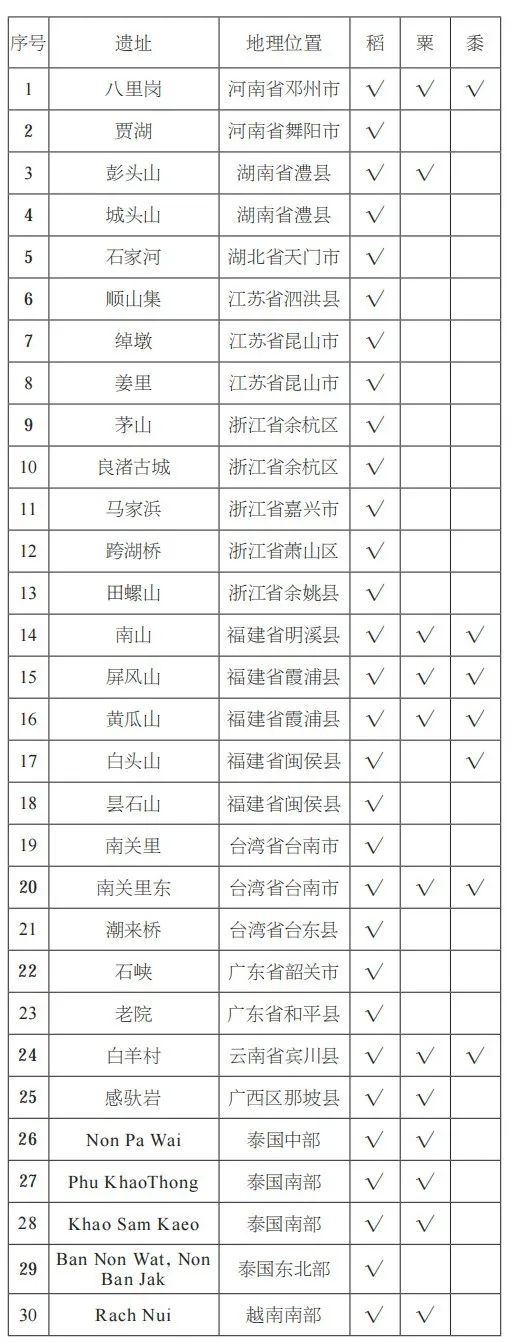
综上所述,最近的研究表明,至少距今4500年前(甚至可早至5000年前),稻米和粟黍类小米在中国东南地区(福建)和台湾地区一出现就已经是共存的。由于农田杂草相关植物考古数据很有限,不好确定这些区域种植的稻属植物是水稻还是雨养稻。不过,由于福建遗址多地处山地丘陵之间,我们推测更可能为雨养稻。无论如何,粟黍类遗存同稻属植物共存,并且在台湾南关里东遗址中出土小米的绝对数量还超过了稻属遗存,这都显示出高地雨养种植系统的重要性。
这些新数据还提供了可信的证据,支持该地区作物从长江中游地区传入的可能路线。长江中游新石器文化中发现更早的稻属和小米类植物遗存共存,传播路线可能是从江西北部和浙江西部进入福建北部,由此连接了中国东南地区与中原内陆区域,避开了尚未发现粟黍农业的长江下游新石器文化区。无论如何,作物由内陆传播至此,对福建沿海已有的海洋文化传统是一种融合和适应。
稻米和小米类植物遗存一同传播到中国南方热带地区,标志着以稻作为主有一定比例粟作的谷物农业在距今4500至4000年间进入了东南亚大陆地区(表二)。东南亚大陆目前最早的种子直接测年数据来自泰国中部Non Pa Wai遗址的粟,年代约距今4400到4200年间。而目前在越南、柬埔寨和泰国等地发现的早期稻属遗存,年代上均未超过距今4000年。
然而,关于这些地区新石器时代和农业定居始于何时仍存在争议,最早的合理估计约为4400年前,最晚的约为4000年前。在越南北部,约4300年前开始出现了具有外来殖民者体质特征的人骨遗存。在越南南部,沿海Rach Nui遗址的出土证据表明,大约在3500年前至3200年前,该地区同时出现稻米和粟,两种作物都被认为是从附近的内陆地区引进的。在铁器时代,泰国南部一些遗址(Khao Sam Kaeo和Phu Khao Thong)除了有来自南亚印度地区的稻米和其他作物,也同时发现了粟,年代大约为距今2400—2000年间。这两处铁器时代泰国遗址的农田杂草数据显示,其种植的稻属植物可能属于雨养稻系统。
在整个东南亚地区,从旱稻到水稻的转变发生在史前时期晚段或历史时期。最近泰国东北部的Ban Non Wot遗址和Non Ban Jak遗址进行的研究提供了一个长程的区域植物考古数据序列,时间跨度从3000年前至1300年前(表二)。在这一时期内,旱稻杂草逐渐减少,大约距今2100年时开始出现水稻伴生杂草;随后水稻杂草逐步增加,到距今1500年时,旱稻杂草消失。这一地区在古气候环境日益干旱的情况下,通过灌溉等措施反而增强了水稻农业的发展,这也说明该地区日益分化的复杂社会能为更加集约化的水稻生产投入更多的劳动力。尽管雨养稻在东南亚的山地一带一直延续到近代,但在大部分平原地区,水稻农业长期以来一直是主要作物经济,供养着该地区历史长河中的城市和国家体系。综上,东南亚大陆地区的水稻种植是在社会复杂化进程和可能的人口增长推动下的农业新发展,而不是早期推动新石器化和区域人口变迁的主要动力。
稻米背后的意义远不止简单的一种“作物”。作为现代作物的稻米,它展现了广泛而多样的生态适应性——从赤道到北纬40度,从接近海平面的低地到海拔超过2000米的山地、高原。遗传学证据显示出现代栽培稻背后存在多个野生种群的影响和不同时期适应和文化选择的多条路径。就像稻米在进入新地区并响应当地野生种群的基因渗入从而在生态上发生了变化一样,移植稻米的农业文化很可能也经历了新的文化适应,并在与当地文化传统的互动之后发生转变,这包括狩猎-渔猎-采集人群和假设的块茎栽培者。这意味着考古学和植物考古学在讨论稻作传播到东北亚和东南亚地区时的挑战,是要理解稻作栽培的出现在当地环境中的历史背景,其中稻米的生态学特征和其在当地生业经济中的地位可能会各不相同。单纯依靠陶器风格和稻米遗存的有无作为指标来推测人群迁徙和稻作农业传播等复杂问题是远远不够的。不同的生计策略,包括多种耕作系统和不同的稻属植物,都会对人口增长、社群分裂和迁移产生不同的影响。
就理解稻作农业出现而言,我们可以区分出三种主要模式。首先,是从本地区野生祖本的利用、栽培进而将其驯化的本土稻作驯化和起源模式,比如中国长江中下游地区。来自长江下游地区的植物考古学数据清楚地说明了这一过程,其中,非落粒性特征的出现演化到占主导地位显示了水稻驯化的完成,并伴之以驯化后的持续演化,包括谷粒大小和比例的变化以及扇形体植硅体的尺寸变化等等。在该地区,伴随着社会复杂化进程,湿地水稻农业推动了人口密度的增加,同时人群与内陆淡水湿地的联系也日益紧密。如前文所说,长江下游水田稻作农业的发展似乎并没有推动人群向外迁移、扩散。
其次,稻米作为已经驯化的作物品种被引进新的地区。这种传播可以通过两种方式实现:一种是被当地人口作为现有生计系统尤其是农业文化中的“附加”品种有选择地采纳。在中国北方中原地区、中国东北及朝鲜半岛、日本列岛等东北亚的广大区域,都有大量稻属植物作为“附加作物”的考古学证据。在这些主要栽培其他驯化谷物(粟黍)的地区,稻属植物作为一种补充作物,成为当地生计策略中的一环。水稻或旱稻的重要程度取决于微环境(如水资源可用性)和社会发展(如劳动力可用性)等条件的好坏,当地人群选择高强度的水稻种植还是低投入低产出的雨养稻系统均受这些因素影响。
值得关注的第三种模式,也是稻作传播的另一种方式,即通过移民携带。也就是说稻米是作为移民文化的一部分得到传播,移民文化包括饮食习惯、生产技术和相应的社会组织。由于水稻种植对劳动力的需求高且具有较高的本地承载能力并不驱动迁徙,所以它不太可能通过这种方式传播。相反,旱稻的属性更适合这样的传播和人群迁徙。这就引发了一个非常关键但尚未解决的问题:旱稻栽培发生于何时、何地,传播过程中又经历了怎样的适应性演化?目前比较清楚的是,传入朝鲜半岛之前,雨养稻似乎已经存在于山东半岛;不太清楚的是,长江中游分布范围广泛背依周围山地位于盆地边缘的新石器晚期城址和聚落采用的是何种稻作种植系统,同共存的占比很低的粟类农业种植体系有何差别;而在长江中下游以南的山地丘陵地带,旱稻种植也许存在多次独立的适应与演化——这些都有可能发生在稻作传播到福建或广东之前或彼时。上述推测和假说突显了系统开展植物考古学研究的重要性,对中国南方和东南亚的植物考古大遗存和植硅体组合进行系统分析与比较研究,才能重建各地区各时期的生业经济体系,理解稻作农业的具体形态和作用。
长期以来,转向稻作农业一直被视为是推动人群迁徙和改变东亚新石器时代社会人口结构的一种“黑箱”机制。然而正如前文所述,生计活动的细节至关重要。事实上,水稻种植体系在新石器时代社会中似乎达到了与人们假设相反的效果,它更有利于支撑当地人口数量和密度的增长,促进和加强对淡水湿地资源的进一步开发利用,进而推动社会分工和社会组织能力,加速区域社会的复杂化进程;并不会因稻作农业发生就迫使人群出现外迁。在长江下游区域的案例中,更不会因为湿地稻作农业发展,促使已经绑定内陆淡水资源的社群面向大海,开始远洋航行。而从水田稻作到雨养稻种植系统的转变,或者整合旱稻和强度更低的雨养谷物小米作物的农业实践,更有可能推动早期农民的向外迁徙和作物的传播。最近数据突显了粟黍类小米在福建、广东、台湾和东南亚大陆新石器农业文化中与稻米共存甚至并重的重要性,也进一步证实了这一观点。
因此,泰国铁器时代出现转向密集型水稻农业的变化也可以置于上述情境中理解,它更有可能是城市化进程的推动因素或者结果,而不是建立新石器时代农业化人口的推力。早期驯化发展的水稻经济具有非扩散的特征,并且在新石器时代旱稻加小米种植系统大规模扩散之前,需要在某些区域已然建立起该种植系统的农业传统,这两点有助于解释水稻栽培(早于距今8000年普遍出现)、水稻驯化(大约距今6000在长江中下游均已完成)和东南亚以谷物为基础的新石器时代开始(晚于距今4500年)之间较长的时间差。基于不同栽培系统所需的生产力、土地承载能力和产量,我们对此提出了一些解释。大量证据表明,比起高产的水田稻作农业,低强度的雨养稻和其他旱作农业更有可能支持新石器时代人口的裂变和向外迁徙。由此可以得出结论:水稻栽培对人口和社会带来的是“向心力”,而低生产力下的旱稻和小米类农业则可视为一种“离心力”,不断推动着人口向外寻找更多的土地。