宁夏隆德县沙塘北塬遗址出土家养黄牛的DNA研究
摘要:运用古DNA技术对宁夏隆德县沙塘北塬遗址2016年发掘出土的家养黄牛骨骼样本进行遗传学分析。共获得8例古代家养黄牛的DNA样本,选择套叠引物对线粒体DNA控制区(285bp)进行PCR扩增。结果显示,这8例个体都属于家养普通牛(Bos taurus),共检测出5个不同的单倍型,归属于2个不同的单倍型类群T3和T4,以T3为主占87.5%,T4次之占12.5%。与其他中国古代家养黄牛的DNA数据进行比较,结果显示沙塘北塬遗址古代家养黄牛与中国甘青地区新石器时代家养黄牛的母系亲缘关系非常接近。本文为了解和认识公元前2000年左右进入中国境内的早期家养黄牛的遗传特征提供了新的证据。
考古学研究表明,公元前4000~前2000年,中国西北地区已经成为东西文化交流的重要区域,一些重要的家养动物(例如黄牛、绵羊、山羊和马等)虽然起源于西亚或中亚,但是很可能在这个时期经由该地区进入中原[1][2][3][4]。对于探索这些家养动物引入中国的路线,揭示从新石器时代向青铜时代过渡时期黄河上游地区不同考古学文化对这些家养动物利用策略的多样化,甘青地区具有至关重要的作用。宁夏南部地区在史前考古学文化区系划分中属甘青地区,旧石器时代晚期至新石器时代的考古遗址发掘证据表明,该地区一直都是东西方文化交流非常活跃的地区。
本文要报道的古代家养黄牛DNA研究的骨骼样本来自于宁夏沙塘北塬遗址,其地理位置为宁夏回族自治区固原市隆德县沙塘镇街道村北侧、渝河北侧二级阶地上。2013年、2015年和2016年宁夏文物考古研究所分别对该遗址进行发掘,发现有房址、灰坑、墓葬、窑址及灰沟,出土遗物丰富,有陶器、石器、骨器、角器等。研究认为该遗址以粟、黍等农业经济生产为主,兼营猪、牛、羊等家畜饲养,也存在着狩猎经济[5][6]。对2013年发掘标本采集进行碳十四年代检测,其年代范围大致为公元前2200年~前1900年之间,属齐家文化早期遗存[7][8]。发掘者最新研究认为以沙塘北塬遗址为代表的文化遗存应该属于常山下层文化向齐家文化的一个过渡类型[9]。
本文系该遗址于2016年发掘出土家养黄牛的古DNA研究。对考古遗址出土黄牛遗存进行了采集、DNA提取,并选择线粒体DNA控制区片段(D-loop)进行PCR扩增,对获得的DNA序列进行线粒体DNA单倍型分型。在此基础上将该遗址发现的家养黄牛与国内外(特别是宁夏地区)的黄牛DNA数据进行比较,以期对新石器时代晚期至青铜时代早期宁夏地区的早期家养黄牛群体遗传特征进行比较研究。
一、材料与方法
(一)样本采集
本文报道的家养黄牛骨骼样本,均来自于沙塘北塬遗址2016年发掘的灰坑或地层中,其中有部分个体来自于相同的遗迹单位。所采集样本骨质较好,利于DNA的提取和随后的实验研究,骨骼保存情况可见图一,样本具体信息见表一。动物考古学家对其进行种属形态鉴定,全部都为家养黄牛。
(二)样本处理与DNA提取
首先使用钻头机械打磨骨骼表面和截面以去除表面尘垢,使用次氯酸钠溶液(有效氯6%)浸泡和紫外照射(UVP,美国,波长254nm),以进一步去除骨骼表面可能存在的外源DNA污染,之后使用液氮冷冻研磨机(SPEX,美国)在液氮环境下将骨骼打磨成粉。在此基础上取0.2~0.5g骨粉,使用Yang等[10]提出的硅柱离心法进行DNA提取。每个样本最终得到约100uL的DNA样本用于PCR扩增与测序。

图一 本文采集的沙塘北塬遗址家养黄牛骨骼情况
(三)DNA扩增与测序
首先,使用引物L1269/H1346[11]扩增线粒体基因组中12S rRNA基因,获取76bp的短片段DNA,对所选样本进行初步种属鉴定;选择Yang等[12]、Troy等[13]和蔡大伟等[14]设计的四对套叠引物来扩增黄牛线粒体DNA控制区的片段,并将这些片段进行连接,共获得黄牛线粒体DNA控制区331bp的序列(16004~16334,包括引物长度)。使用AmpliTaq GoldTM聚合酶(Thermo fisher,美国)进行PCR扩增(60个循环),具体PCR反应体系与扩增程序设置见参考文献[15]。扩增产物用2%琼脂糖凝胶进行电泳检测。PCR阳性产物送到测序公司直接测序,正反引物双向测序。基于本课题组在古代牛骨DNA的实验经验[16][17],在本实验的PCR扩增过程中同样未使用BSA(牛血清白蛋白)。
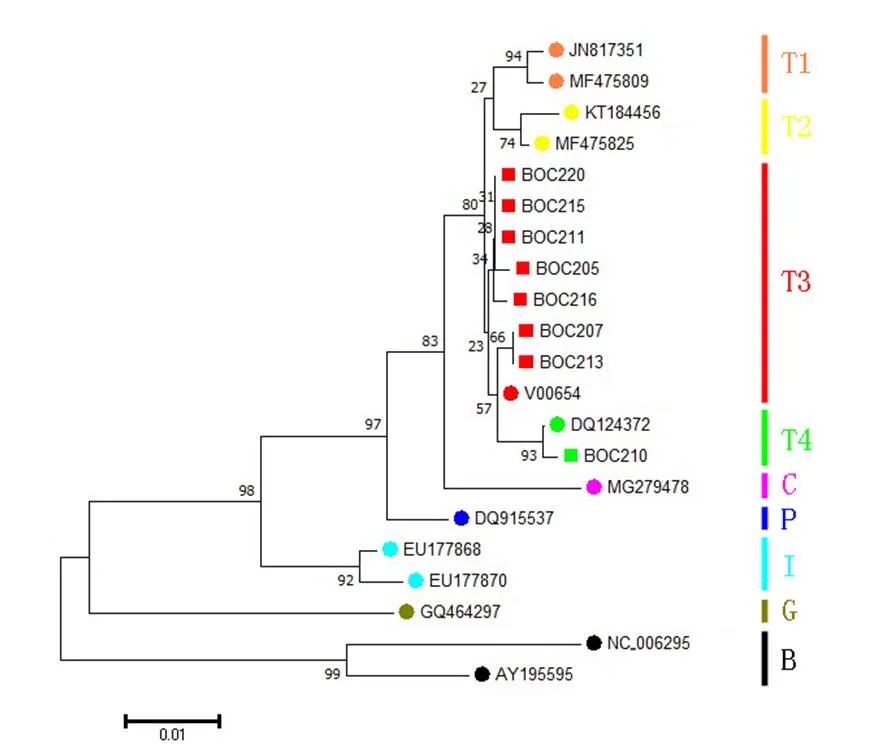
图二 系统发育邻接树
注:本文研究样本用方块表示,用BOC标注编号。对比序列来自GenBank数据库,用圆圈表示,序列编号为GenBank号,包括:普通牛单倍型类群T1(GenBank:JN817351和GenBank:MF475809)、普通牛单倍型类群T2(GenBank:KT184456和GenBank:MF475825)、普通牛单倍型类群T3(GenBank:V00654)、普通牛单倍型类群T4(GenBank:DQ124372)、瘤牛单倍型类群I(GenBank:EU177868和GenBank:EU177870)、亚洲原始牛单倍型类群C(用C表示,GenBank:MG279478)、欧洲原始牛(用P表示,GenBank:DQ915537)、牦牛(用G表示,GenBank:GQ464297)和水牛(用B表示,GenBank:NC_006295和GenBank:AY195595)。全部节点都标注自展值,比例尺表示每一个位点的核苷酸替代率。
(四)数据分析
使用Chromas Pro软件对获得的DNA序列进行序列信号的读取和检测,使用Clustal X2软件[18]对DNA序列进行比对,使用BioEdit软件[19]进行编辑和连接。使用MEGA 7软件[20]构建系统发育树,详见图二。使用Dna-SP(v.6.12.03)软件[21]和Arlequin(v. 3.5)软件[22]计算单倍型多样性与核苷酸多样性。使用PopART(v. 1.7)软件[23]构建中介网络图(图三),选择中国考古遗址出土家养普通牛作为对比序列,包括:山西陶寺、山西周家庄、河南花地嘴、河南望京楼、河南二里头、河南殷墟孝民屯、陕西泉护村、陕西石峁、吉林后套木嘎、内蒙古大山前、青海长宁、宁夏打石沟、宁夏王大户、宁夏中庄、宁夏九龙山、新疆小河等[24][25][26][27][28][29][30][31][32]。
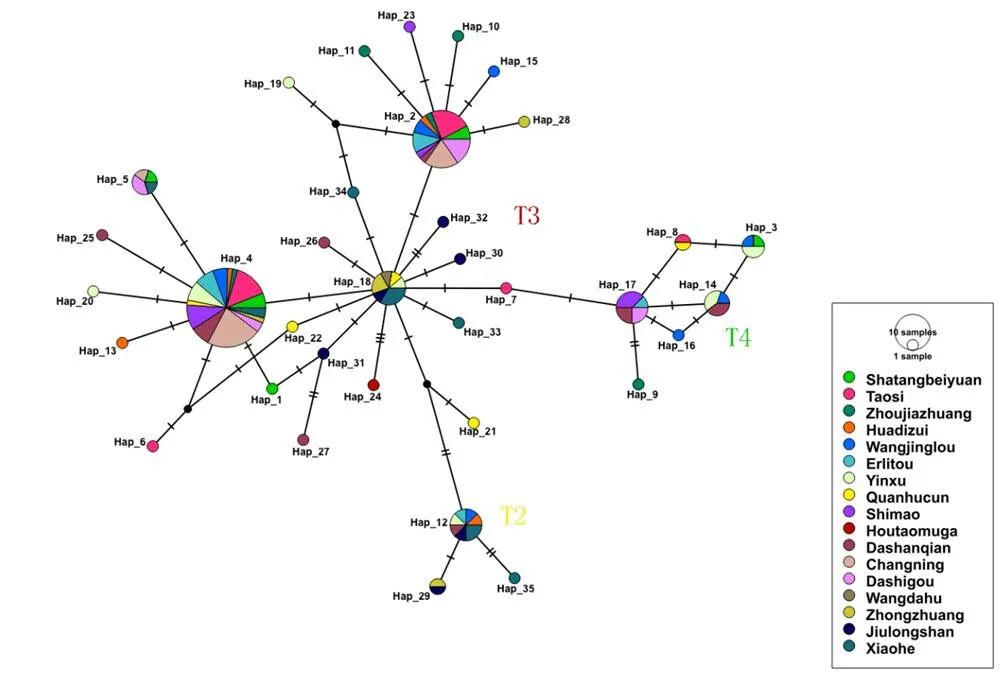
图三 中介网络图
注:每一个圆圈代表一个单倍型,圆圈的大小与共享这一单倍型的样本数量成正比,不同颜色代表不同遗址。
(五)污染控制
全部实验都在专门的古DNA实验室进行,严格按照防污染措施操作,具体实验操作详见[33],以最大限度地防止外源DNA污染的可能。所有的DNA序列都得到了正反引物双向测序结果和重复,用以识别和排除实验中可能存在的误差。所有的DNA提取和PCR扩增中都加入了空白对照,用以识别和排除实验室系统和试剂、耗材等可能存在的污染。
二、结果
本文全部实验的DNA提取和PCR扩增的空白对照都为阴性,未检测到实验污染,并且所获得的DNA序列都得到了重复验证,显示出所获得的DNA数据的可信性。
在采集的8例样本中,全部获得了线粒体基因组中12S rRNA基因76bp的部分DNA序列,在GenBank数据库中(http://www.ncbi.nlm.nih.gov)对这些DNA序列进行了BLAST共享序列搜索,结果发现全部样本的共享序列都属于牛属(Bos)。这8例样本都获得了线粒体DNA控制区的285bp序列(16029~16313,去除两端引物),在GenBank数据库中(http://www.ncbi.nlm.nih.gov)对这些DNA序列进行了BLAST共享序列搜索,结果发现它们都属于家养普通牛(Bos taurus)。
以V00654[34]作为参考序列,将这8个线粒体DNA控制区序列与之进行比对,实验结果见表二。共检测出8个多态性位点,全部为转换,没有颠换、插入或缺失,转换在嘧啶之间的比例略高占75%(6/8),遗传多态性不高。在这8个序列中共检测出5个单倍型,BOC211、BOC215和BOC220, BOC207和BOC213分别共享有相同的单倍型。根据家养普通牛单倍型类群划分的已有研究成果[35][36][37][38],基于线粒体DNA控制区序列变异模式,本文发现的5个单倍型可以归属于2个不同的单倍型类群T3和T4,以T3为主占87.5%,T4仅发现一个个体(BOC210)占12.5%,未见其他单倍型类群。
三、讨论
目前已发表的关于中国古代家养黄牛DNA研究的材料主要分布在中国北方地区,年代范围从新石器时代至青铜时代。研究结果显示这些家养黄牛都属于普通牛(Bos taurus)的线粒体DNA单倍型类群[39][40][41][42][43][44][45][46][47],并且在公元前2000年左右单倍型类群T2、T3和T4在中国已经全部出现[48]。从地理分布情况来看,这些研究所涉及的遗址主要分布于中原地区和甘青地区。从年代来看,属于新石器时代的材料非常少,仅有吉林后套木嘎、山西陶寺、山西周家庄、宁夏打石沟、青海长宁几个遗址可能涉及到新石器时代的遗存。
经过对沙塘北塬遗址多次发掘后,发掘者最新研究认为沙塘北塬遗址为代表的文化遗存应该属于常山下层文化向齐家文化的一个过渡类型[49]。结合2013年发掘标本的测年结果(年代范围大致为公元前2200年~前1900年之间)[50],在探讨中国古代家养黄牛时,应该说沙塘北塬遗址是一个非常重要的遗址点。该遗址出土的家养黄牛遗存可能代表着从境外引入中国的一批早期家养黄牛。
本文研究结果显示,沙塘北塬遗址出土的家养黄牛遗存全部属于普通牛的线粒体DNA单倍型类群,可以归属于单倍型类群T3和T4,以T3为主,T4次之(图二),其分布频率与已发表的其他中国古代早期家养黄牛比较相似(图三)。具体地讲,与沙塘北塬遗址家养黄牛遗传结构最为相似的遗址主要包括山西陶寺(龙山时期)、山西周家庄(龙山时期)、陕西泉护村(青铜时代)、陕西石峁(龙山末期至二里头时期)和宁夏打石沟(新石器时代),它们都是同时存在T3和T4,且以T3为主的遗传结构特征。并且在这些遗址中,除了泉护村外,其他几个遗址的年代都可以早到新石器时代。除了上述遗址外,年代能够早到新石器时代的遗址还有吉林后套木嘎(新石器时代),但是该遗址仅发现1例家养黄牛(属于单倍型类群T3),并不具备单倍型类群分布频率的统计学意义。总地来说,在新石器时代,中国家养黄牛的遗传结构相对简单。
从图三中可以看出,在沙塘北塬遗址发现的5个单倍型(Hap_1~Hap_5)中,有4个单倍型(Hap_2~Hap_5)在其他中国考古遗址的黄牛中都有发现。并且其中3个单倍型(Hap_2、Hap_4和Hap_5)同时在位于甘青地区的宁夏打石沟、青海长宁中有发现。结合上述讨论认为沙塘北塬遗址与打石沟的家养黄牛遗传结构具有一致性。打石沟遗址位于宁夏彭阳县古城镇店洼村水库东50米的坡地上,发掘者研究认为该遗址的文化特征属于新石器时代晚期客省庄二期文化(距今4300~4000年)[51]。本文所研究的沙塘北塬遗址所处地理位置与打石沟较近,关于该文化遗存的最新研究认为应该属于常山下层文化向齐家文化的一个过渡类型[52]。二者年代相近,虽然所属文化特征不同,但是依据两个遗址出土家养黄牛的遗传结构特征推测新石器时代甘青地区家养黄牛的母系亲缘关系非常近,即其母系来源可能相同或相近。沙塘北塬遗址中另1个单倍型Hap_3属于T4,在河南望京楼(二里头、二里冈文化时期)、殷墟遗址(商代晚期)的黄牛中找到共享相同序列的样本,望京楼、殷墟的年代都晚于沙塘北塬,不排除甘青地区与中原地区存在交流。结合遗址年代早晚关系,DNA证据显示从西亚经中国甘青地区传入中原,有可能是家养黄牛传入中国的一条重要通道。
就宁夏地区而言,蔡大伟等[53]于2018年报道了采集自宁夏的四个考古遗址(新石器时代晚期的打石沟遗址和春秋战国时期的王大户遗址、中庄遗址、九龙山遗址)的27例古代家养黄牛的DNA研究成果,早期的打石沟遗址家养黄牛只有两种单倍型类群T3和T4,而晚期的王大户遗址、中庄遗址、九龙山遗址则出现了新的单倍型类群T2,表明进入青铜时代以后,宁夏地区的家养黄牛的遗传结构发生了改变。由于从新石器时代到春秋战国时期中间还存在一个很长时间的缺环,所以单倍型类群T2最早何时进入宁夏地区目前尚不清楚,期待有更多该地区的研究成果公布与发表,可以对这一问题作更合理的解释。结合本文的研究结果仍然可以看出宁夏地区一直是家养黄牛传入中原的一个重要通道,并且传入过程是持续不断的。
陈相龙等[54]对沙塘北塬遗址出土人骨与动物骨骼开展了碳氮稳定同位素食性分析。在本文研究的8个个体中,线粒体DNA结果显示都为家养普通牛,其中有7个进行了碳氮稳定同位素分析。它们的δ13C值分布范围与平均值分别是-18.5‰~-15.3‰与-16.5±1.0‰,δ15N值分布范围与平均值分别是7.4‰~10.5‰与9.5±1.0‰,呈现出C3/C4混合型食物结构。综合古DNA与碳氮稳定同位素结果可以看出,该遗址的家养黄牛可能主要以采食C3植物为生,同时也食用了少量粟和黍的秸秆、谷糠等副产品[55]。结合已发表过的动物考古学和食性分析数据,可以认为公元前2000年左右在中国甘青地区多个遗址发现的牛、绵羊和山羊遗存的数量明显增加,说明在新石器时代晚期至青铜器时代早期,以食草动物为主(主要包括牛、绵羊和山羊等)的畜牧业在甘青地区得到了发展[56][57][58][59][60]。
四、结论
通过对宁夏隆德县沙塘北塬遗址出土家养黄牛进行线粒体DNA控制区序列的分析,并结合该遗址所处的重要年代和地理位置,本文得出以下认识:沙塘北塬遗址出土的家养黄牛骨骼遗存全部属于普通牛,可以归属于单倍型类群T3和T4,以T3为主,T4次之,其分布频率与已发表的其他中国古代早期家养黄牛比较相似,并且与同位于宁夏的新石器时代遗址打石沟的家养黄牛遗传结构具有一致性。沙塘北塬遗址古代家养黄牛DNA研究再次证实,在新石器时代中国家养黄牛的遗传结构相对简单。
此外,本文推测以沙塘北塬、打石沟、长宁等为代表的公元前2000年左右甘青地区的家养黄牛还处于家养黄牛引入中国境内的早期阶段。但是随着时间推移和对黄牛需求的增加,以及中原地区大量需求的向外扩散,饲养黄牛的数量和规模逐步发展壮大,新的品种也可能会从西亚、中亚等地区得到不断地引进。
致谢:感谢宁夏文物考古研究所为本研究提供了宝贵的样本,感谢编辑部老师提出的宝贵意见。

注释
(作者:赵欣 中国社会科学院考古研究所科技考古中心 西蒙菲莎大学(Simon Fraser University)考古学系古DNA实验室,东晓玲 刘铭 中国社会科学院考古研究所科技考古中心,张桦 西蒙菲莎大学(Simon Fraser University)考古学系古DNA实验室,侯富任 杨剑 王晓阳 宁夏回族自治区文物考古研究所,陈相龙 张雅军 中国社会科学院考古研究所科技考古中心,杨东亚 中国社会科学院考古研究所科技考古中心 西蒙菲莎大学(Simon Fraser University)考古学系古DNA实验室;原文刊于《南方文物》2021年第5期)
